Transmission de l’influx nerveux
La transmission de l’influx nerveux (IN) est assurée par les neurones (fig A); chaque neurone présente un pôle récepteur fait de prolongements arborescents plus ou moins fournis (les dendrites -D-), un corps cellulaire (CCN) qui contient le noyau (N) et un pôle effecteur représenté par l’axone (A) et ses prolongements axonaux (Pa) qui s’articulent avec le ou les neurones suivants sur ses dendrites et/ou souvent sur le corps cellulaire lui-même. Ces jonctions sont appelées synapses (SY) et constituent l’essentiel du mécanisme de transmission de l’IN qui ne se transmet que dans le sens prolongements axonaux – dendrites.
La naissance et la propagation de l’IN sont basées sur des processus électro-chimiques très complexes dont nous ne présentons ici que les grandes lignes.
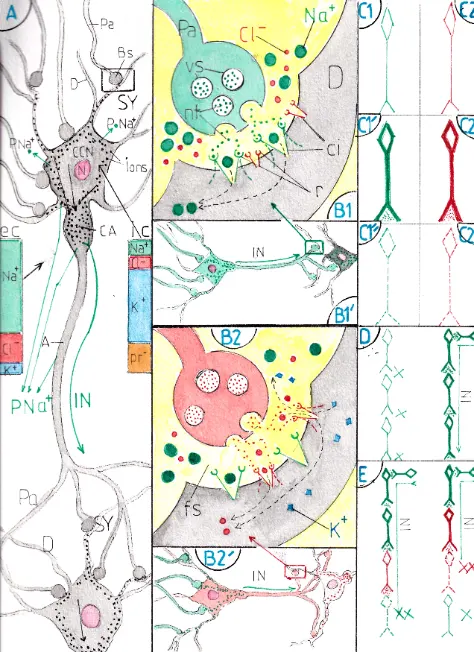
Un neurone au repos est par définition non excité; son maintien au repos est assuré par la polarisation négative de son contenu intra-cellulaire du fait de la présence d’ions négatifs en plus grande quantité que d’ions positif; cette résultante négative est généralisée à la totalité du neurone, dendrites et prolongements axonaux compris; les trois ions principaux (sodium Na+ , chlore Cl- et potassium K+) sont en répartition inégale de chaque côté de la membrane externe du neurone (voir les deux barres colorées de chaque côté de la fig A) : la somme électrique de ces deux compartiments est cependant identique grâce là la présence de protéines chargées négativement (pr-) et non diffusibles du côté intracellulaire. L’espace intracellulaire du neurone serait donc neutre si la membrane cellulaire du neurone n’était pas équipée à sa face profonde de multiples « pompes à Na+ » (Pna+) qui puisent leur énergie dans le métabolisme cellulaire pour éjecter en permanence des ions Na+ dans le milieu extracellulaire et donc maintenir dans la cellule une négativité électrique par rapport à l’espace extérieur.
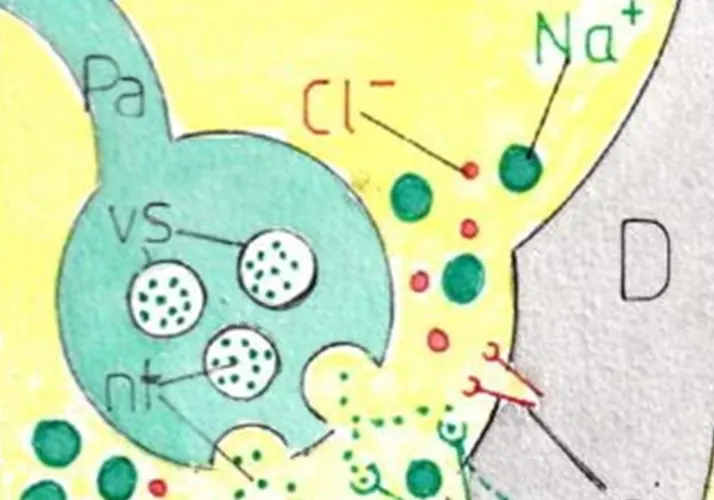
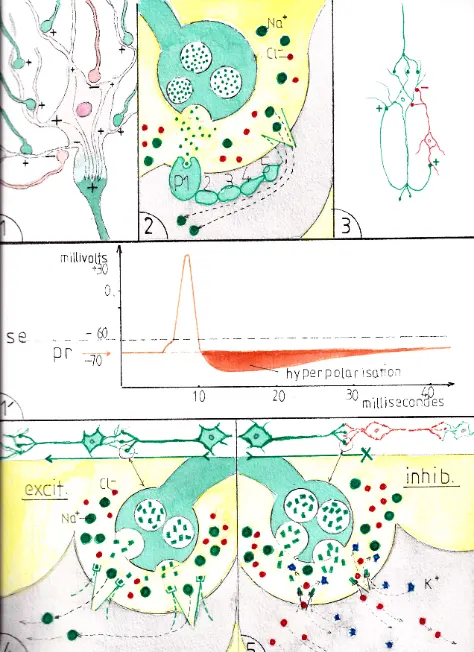
L’IN nait quand des ions positifs (ici les ions Na+) rentrent à nouveau dans la cellule ce qui est facilité par leur très grande abondance dans le milieu extracellulaire : cette entrée est provoquée par l’ouverture des pores (ou canaux ioniques) du versant dendritique de la synapse (SY); le bouton synaptique du neurone d’amont (fig B1) est en effet rempli de vésicules (vs) contenant un neurotransmetteur (nt). L’influx nerveux les poussent à s’ouvrir dans la fente synaptique ; les neurotransmetteurs expulsés se fixent alors sur des récepteurs (r) entourant les canaux ioniques (CI) qui leurs sont spécifiques à la surface des dendrites ; les canaux ioniques concernés s’ouvrent alors ce qui permet le passage des ions adaptés à la dimension des canaux (ici les ions Na+).
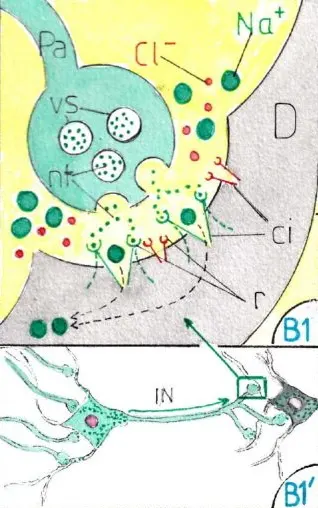
On connait environ 100 neurotransmetteurs et chaque neurone n’en produit qu’une seule variété; par contre des récepteurs différents existent sur les versants dendritiques d’un même neurone; ce qui explique que ce même neurone puisse réagir de façon très différente selon le neurotransmetteur fourni par le neurone d’amont; cela permet de comprendre l’opposition entre neurone excitateur et neurone inhibiteur (fig B1 B’1 et B2 B’2).
Un neurone excitateur expulse par ses boutons synaptiques (fig B1 et B’1) un neurotransmetteur qui va se fixer sur un récepteur gardien d’un canal ionique suffisamment large pour laisser passer les ions Na+ du milieu extracellulaire (pointillés verts); poussés par leur forte concentration extérieure ces derniers s’engouffrent dans tous les canaux ioniques correspondants ; ils rejoignent d’abord le corps cellulaire du neurone; puis (revenir à la fig A) ils s’accumulent à la partie initiale de l’axone dans une zone un peu élargie appelé cône axonal (CA) : lorsque la somme des ions Na+ a suffisamment dépolarisé ce cône initialement négatif, elle déclenche , au sein de ce cône un potentiel de dépolarisation qui va se poursuivre tout le long de l’axone jusque qu’aux prolongements axonaux; les vésicules synaptiques des boutons suivants vont s’ouvrir et ainsi prolonger la transmission de neurone en neurone si ces derniers sont également excitateurs; ces neurones sont dits excitateurs du fait de leur capacité à délivrer pour le (ou les) neurone suivant un neuro transmetteur à pouvoir excitateur.
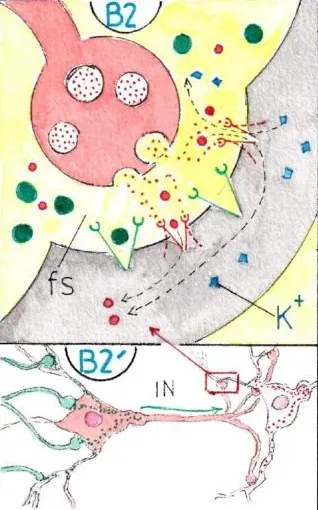
Un neurone inhibiteur est un neurone qui bloque au contraire l’influx nerveux; il est représenté en rose sur la fig B2′; dans ce cas ses boutons synaptiques (fig B2) sont remplis de neurotransmetteurs chimiquement différents; lorsque les vésicules qui les contiennent s’ouvrent dans la fente synaptique les neurotransmetteurs viennent se fixer sur des récepteurs différents (ici en rouge) entourant des canaux ioniques plus étroits, ne laissant passer que des petits ions : du fait des différences de concentration entre les milieux intra et extra -cellulaires , les ions potassium (K+) s’échappent de la dendrite et les ions chlore (CL-) y entrent ce qui amène un surplus d’ions négatifs dans le neurone d’aval; celui-ci s’hyperpolarise (négativement) et ne peut plus transmettre d’influx nerveux : on le dit inhibé. Un neurone inhibé (hyperpolarisé) est différent électriquement d’un neurone au repos : il est en effet davantage polarisé négativement et pour être remis en activité il devra être davantage excité qu’un neurone au repos.
Bien comprendre que le neurone inhibiteur est différent du neurone inhibé; il a du être au contraire être excité par un neurone excitateur pour entrer en action et délivrer ses neurotransmetteurs inhibiteurs !
Les 6 fig C montrent 3 états différents de neurones excitateurs (en vert) et inhibiteurs (en rouge) : en traits fins les neurones sont au repos (C1 et C2); en traits épais ils sont en activité et ils sécrètent leur neurotransmetteur (C1′ et C2′ ) ; en pointillés ils sont inhibés (C1 » et C2 »).
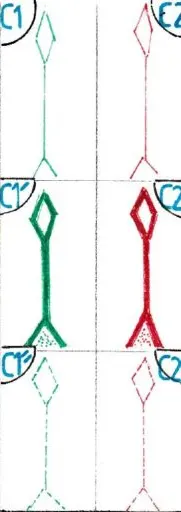
La fig D montre : à gauche 4 neurones excitateurs au repos , sans sécrétion de neurotransmetteur, donc sans passage d’IN; à droite un neurone excitateur lui même excité a permis le démarrage de l’IN sur l’ensemble de la chaine.
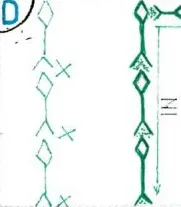
Sur la fig E : à gauche est interposé un neurone inhibiteur dont l’excitation a hyperpolarisé donc bloqué l’IN sur le dernier neurone ; à droite un montage plus complexe mais fréquent dans le système nerveux central est celui de la présence de deux neurones inhibiteurs successifs : celui d’amont en inhibant celui d’aval empêche ce dernier d’inhiber le neurone excitateur suivant et donc le libère en partie ; certes il ne conduit pas momentanément d’IN , mais à l’inverse de son homologue de gauche il lui suffira d’une faible excitation pour redémarrer.
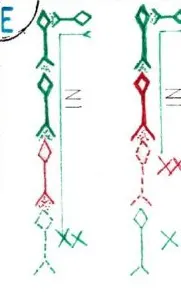
Dans la réalité la transmission de l’IN ne doit pas être envisagée de façon parcellaire comme ici , mais globale ; en effet quand on sait qu’un neurone est en général connecté avec de 10 à 1000 autres neurones et inversement et que chaque connexion se fait avec un nombre très élevé de synapses , il est impossible de suivre le passage de l’IN d’un neurone à l’autre ; il faut se contenter , et c’est déjà beaucoup, de pouvoir circonscrire des régions à pouvoir excitateur ou inhibiteur prépondérants et d’autres à pouvoir plutôt modulateur.
Ce pouvoir modulateur est souvent du à des connexions voisines entre neurones excitateurs et inhibiteurs établissant pour les neurones connectés la possibilité de basculer dans l’une ou l’autre des situations suivant les influx eux-mêmes excitateurs ou inhibiteurs de voisinage; en effet la plupart des neurones possèdent à la fois des récepteurs excitateurs et inhibiteurs . Ainsi le neurone de la fig 1 est soumis à un certain nombre de stimulations excitatrices et inhibitrices du fait de connexions axonales venant de neurones différents.
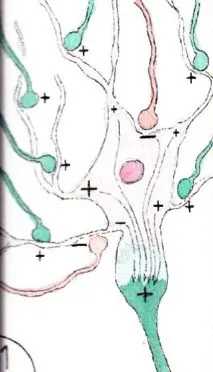
Par ailleurs plus un bouton synaptique se trouve prés du corps cellulaire du neurone ,moins il y a de déperdition de charges électriques , lesquelles diminuent en effet le long des dendrites dont la membrane est en partie perméable aux charges électriques de voisinage ; si bien que la somme de ces charges qui se rassemblent sur le cône axonal peuvent varier en fonction de l’activité des neurones d’amont ; sur le schéma 1 la somme est positive et le neurone se trouve excité.
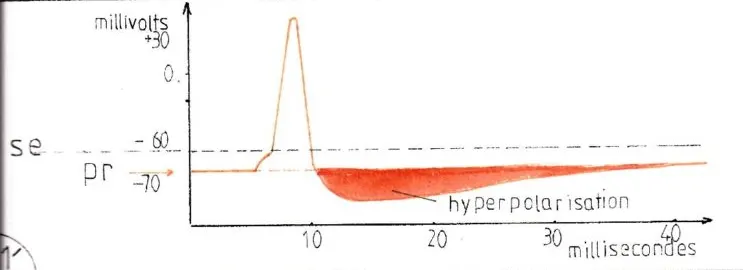
La fig 1′ représente le graphique classique de l’évolution du potentiel d’action d’un neurone excité en fonction du temps (le potentiel de repos se situant autour de – 70 millivolts ); la petite montée initiale correspond à l’accumulation des charges ioniques positives dans le corps cellulaire du neurone jusqu’à – 60mv qui correspond au seuil excitateur (se) ; à ce moment le cône axonal qui a recueilli la somme électrique se trouve à +10 mv (en effet la répartition homogène des charges électriques dans l’ensemble du neurone ne se fait pas immédiatement ) ce qui entraine l’ouverture massive des canaux ioniques membranaires du neurone et provoque le potentiel d’action qui va se propager tout le long de l’axone ; cette période ne dure habituellement que quelques millisecondes et comprend pourtant toutes les phases de la transmission synaptique ; pendant la phase ascendante et descendante du potentiel d’action jusqu’au retour à -60 mv le neurone reste hyperexcitable et de nouveaux potentiels d’action peuvent s’enchainer de façon subintrante jusqu’à pouvoir réaliser un état tétanique ; par contre lorsque le potentiel d’action se termine le neurone s’hyperpolarise jusqu’à -90mv : s’ensuit une période réfractaire pendant laquelle le neurone n’est plus excitable sauf par un afflux important de synapses excitatrices.
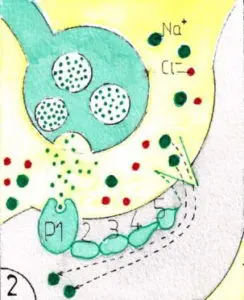
Certaines synapses font intervenir des protéines réceptrices intermédiaires entre les neurotransmetteurs et l’ouverture des canaux ioniques (fig 2) ; elles sont appelées métabotropes ; cette chaine protéique peut être plus ou moins complexe (ex 1,2,3, 4 et 5 du schéma) ; on comprend facilement que la transmission synaptique de ce type puisse être beaucoup plus longue en durée que quelques millisecondes et atteindre la seconde , la minute , voire l’heure , multipliant ainsi à l’infini les variétés de transmission de l’IN.
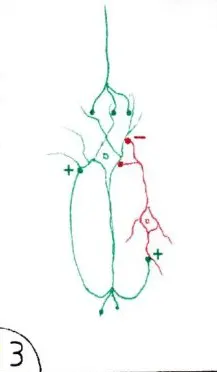
La voie de la rétroaction (fig 3) constitue aussi une possibilité de prolonger une action excitatrice lorsqu’un neurone envoie des prolongements axonaux sur ses propres dendrites ou son propre corps cellulaire (en vert)… ou d’arrêter son excitation en transmettant l’influx à un neurone inhibiteur (en rouge) qui se réfléchit sur lui.
Signalons aussi l’hypersensibilité à l’excitation des neurones excitateurs ayant été hyper-stimulés peu de temps auparavant.
A l’inverse et plus logiquement des neurones hyper-stimulés récemment peuvent ne plus pouvoir exciter les neurones suivants en raison d’un épuisement synaptique : en effet on estime qu’un bouton synaptique ne peut guère fournir plus de 1000 neurotransmissions à la suite par épuisement de ses réserves en neurotransmetteurs ; ceux-ci devront être reformés dans le corps cellulaire du neurone et redescendre le long des travées axonales avant que le neurone ne puisse redevenir fonctionnel.
En effet si le système nerveux central est en perpétuelle activité les différentes parties doivent récupérer à tour de rôle ; ces interruptions sont absolument nécessaires et utilisent surement les deux voies précitées que sont l’épuisement synaptique et le système neuronal inhibiteur.
Par ailleurs de passage de l’IN par les processus physico-chimiques de la synapse semble entrainer également des conséquences plus complexes dans la « machinerie » cellulaire du neurone , comme le montrent des études récentes ; en effet les ouvertures des canaux ioniques peuvent provoquer des chaines de réactions moléculaires au sein de la cellule , mais également de son noyau jusqu’à atteindre des régulateurs d’expression de l’ADN qui favoriseraient une excitabilité acquise de certains neurones lors de nouvelles stimulations ; ce processus a été étudié chez l’animal au niveau d’une structure très particulière du cerveau commune avec l’homme, l’hippocampe responsable de la mémoire à long terme ; cette excitabilité pourrait de fait se maintenir dans le temps et permettre un début de compréhension de cette mémoire ; de même qu’un mécanisme inverse, lui aussi actif , pourrait favoriser l’oubli sans lequel les processus mnésiques seraient vite totalement débordés.
La courte liste de certaines de ces modulations permet de se faire néanmoins une idée du fonctionnement infiniment complexe du système nerveux central et de la difficulté à établir des relations absolues entre ses différentes parties et fonctions; les neurotransmetteurs eux-mêmes peuvent avoir dans certains cas des actions opposées suivant les récepteurs sur lesquels ils agissent :
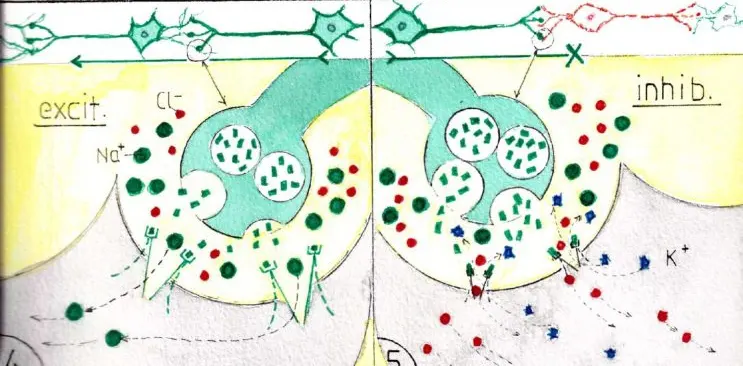
Les fig 4 et 5 montrent un exemple d’un même neurotransmetteur ayant des actions excitatrices ou inhibitrices selon les récepteurs qui les reçoivent : en 4 les nt sont compatibles avec des récepteurs de canaux ioniques favorisant le passage des ions Na + : les neurotransmetteurs seront donc excitateurs pour le neurone suivant . En 5 les nt et les récepteurs sont les mêmes mais ces derniers sont gardiens de portes plus étroites ne permettant le passage que des ions Cl- et K+, les mêmes nt devenant donc inhibiteurs pour le neurone suivant. Ceci explique pourquoi certains neurotransmetteurs peuvent avoir des effets très différents selon les récepteurs des neurones avec lesquels ils sont connectés. En effet si un neurone a habituellement plusieurs types de récepteurs il n’a pas tous les types et ceux qu’il possède le caractérise et le rend réceptif à un nombre limité de neurorécepteurs .
Pour terminer indiquons quelques neurotransmetteurs parmi les plus souvent cités en physiologie du système nerveux ; mais plus de 100 sont actuellement connus !
- L’acétylcholine: le plus souvent excitatrice
- La noradrénaline : inhibitrice ou excitatrice suivant les cas
- La dopamine :idem
- La glycine et l’acide gamma -amino-butyrique (GABA) inhibiteurs
- L’acide glutamique : excitateur
- La sérotonine :excitateur
- L’histamine :excitateur
Ce résumé n’a pas la prétention d’ être un cours sur le fonctionnement de l’influx nerveux mais il peut aider des non spécialistes à mieux appréhender certains passages de la physiologie des différentes fonctions du SNC exposées plus loin . Pour les lecteurs qui seraient passionnés par le sujet on ne peut que recommander la bible des Neurosciences : le classique Purves remis à jour au fil des nouvelles éditions… Mais dont l’accès reste difficile pour les non avertis.
PS : les couleurs vertes et rouges utilisées sur les schémas de cette page ne concernent qu’elle ; pour les pages suivantes elles peuvent n’avoir aucun rapport avec le caractère excitateur ou inhibiteur des faisceaux nerveux sauf pour les indications en + ou – qui restent valables ; en effet pour la suite il est classique de représenter les fibres motrices en rouge et les fibres sensitives ou sensorielles en bleu.