Olfaction et Goût
A – L’olfaction ou odorat
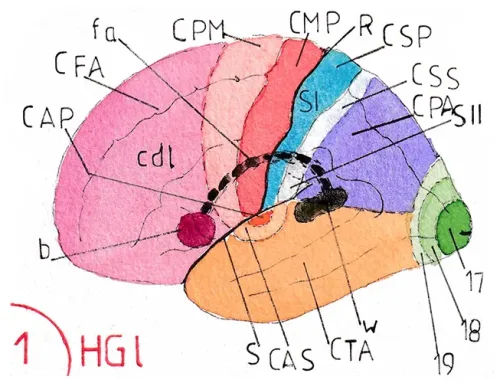
Moins développé que chez beaucoup d’espèces animales chez lesquelles elle joue un rôle vital, l’olfaction humaine nait de l’épithélium olfactif situé à la partie supérieure des fosses nasales (zone bleue très claire, presque blanche) ; il tapisse le versant latéral de chaque fosse ; soit le cornet supérieur (cs) et la partie supérieure du cornet moyen (cm) – fig1–

Sur le versant médial il ne recouvre qu’une partie réduite de la cloison nasale (cn) – fig2 –


Les cellules sensorielles (fig3) qui transmettent les influx olfactifs sont inclues dans l’épithélium ; leurs extrémités sont pourvues de cils récepteurs qui se prolongent dans une couche de mucus (M) sécrétée par des glandes muqueuses intégrées à l’épithélium ; les axones traversent la lame criblée de l’os ethmoïde (LCE) pour rejoindre le bulbe olfactif –BO– (un de chaque côté) ; cette traversée explique le risque de lésions de ces axones dans les traumatismes crâniens antérieurs en cas de fracture de cet os.

La fig 4 montre à un grossissement plus important des éléments encore plus précis : d’abord la présence autour des cellules olfactives (co) de cellules de soutien (cs) et de cellules basales (cb), ces dernières étant chargées de remplacer les cellules olfactives à la fin de leur cycle qui dure environ 1 mois, propriété de renouvellement qu’elles partagent avec les cellules gustatives .
Les stimuli chimiques portés par l’air ambiant et par ceux issus du carrefour oro-naso pharyngé (et venant compléter les sensations gustatives) sont très mal connus et surtout sans doute extrêmement nombreux : cependant certaines odeurs caractéristiques peuvent être testées dans les troubles de l’odorat : une cinquantaine sont assez classiques mais il n’y a pas de liste limitative comme dans d’autres explorations sensorielles.
Les axones des cellules olfactives se regroupent en bouquet pour traverser la lame criblée, les perforations osseuses étant en nombre limité ; elles se dirigent vers des formations arrondies situées à la face profonde du BO et portant le nom de glomérules olfactifs (go) ; à ce niveau elles entrent en connexion avec les dendrites de cellules dite mitrales (cmit) dans une proportion de 1 à 1000 ; chaque glomérule comprend les synapses de 25000 cellules olfactives pour 25 cellules mitrales. Quand un glomérule est actif il inhibe en général les glomérules adjacents par des cellules péri-ganglionnaires (cpg) ; et plus haut les cellules mitrales en activité reçoivent des influx inhibiteurs des cellules granulaires (cgr) ; elles même sont sous l’influence d’influx descendant (en rouge pointillé) venant du tractus olfactif (TO) ; toutes ces inhibitions sont sans doute utiles pour éviter la saturation des centres olfactifs supérieurs du fait de la dispersion intranasale des stimuli olfactifs.
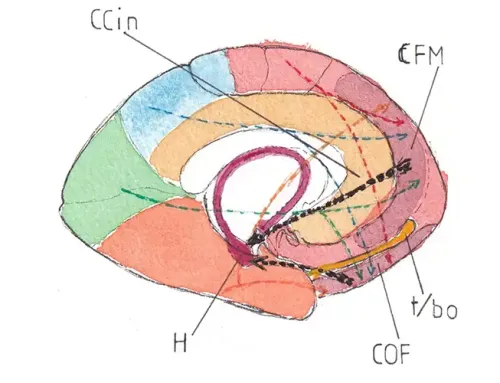
Les axones des cellules mitrales se répartissent en deux voies après leur sortie du tractus optique (TO) –fig3 et fig 5 -: la voie médiane se dirigeant vers la commissure antérieure (CA) et de là vers l’hypothalamus sans doute responsable des réactions réflexe de salivation et même d’éveil chez un sujet endormi ; elle aurait aussi une fonction inhibitrice sur le bulbe controlatéral peut-être pour aider à localiser l’origine d’une odeur. La voie latérale se termine dans le lobe piriforme (LPT) du cortex temporal antérieur où elle rejoint le partie subcorticale de l’amygdale (A) juste en dehors de l’uncus (U) mais aussi la partie postérieure du cortex orbito- frontal (PCOF) responsable de la discrimination olfactive.
La fig 5 montre une vue de la face inférieure du cerveau et de la position du bulbe et du tractus olfactifs et des terminaisons corticales.

NB : pour faciliter les repérages : LT et LF : faces inférieures des lobes temporal et frontal ; CHO : chiasma optique ; H : hypophyse ; TC : coupe horizontale du tronc cérébral.
De ces deux régions principales les sensations olfactives atteignent des régions secondaires comme l’hippocampe (mémoire olfactive) et le cortex cingulaire (réactions affectives diverses).
A préciser qu’il n’y a pas vraiment de nerf olfactif au sens habituel du mot ; le nerf correspond uniquement aux axones des cellules olfactives traversant la lame criblée ; le nerf olfactif pris au sens large (bulbe et tractus optique) ne traverse pas la ligne médiane comme les autres nerfs crâniens.
B- Le goût
Les cellules réceptrices du goût (cellules gustatives) se trouvent essentiellement sur la langue mais il en existe également sur le palais, voire plus en arrière au niveau de l’épiglotte (e) et de la région naso- pharyngée (rnp) – fig 1–


L’essentiel est cependant lingual, notamment dans les sillons qui entourent les papilles caliciformes (pc) fig2 marquant le V lingual, limite anatomique classique entre partie antérieure et partie postérieure de la langue ; ces deux parties sont assez riches elles aussi en récepteurs du goût formés de cellules apparemment identiques mais pouvant présenter des sensibilités différentes aux stimuli : ainsi les cellules réceptrices du V lingual seraient plus sensibles à la saveur amère, celles des bords de langue au salé et à l’acide et les antérieures au sucré. On ajoute actuellement l’unami (le délicieux !) correspondant aux acides aminés aromatiques, notamment ceux contenus dans les aliments riches en protéines.
Cependant l’étude du goût ne serait pas complète si on ne faisait pas intervenir 3 éléments complémentaires :
- La volatilité des odeurs des aliments qui parviennent à la muqueuse olfactive par les narines et par les choanes (ch) fig 1
- La nécessité de libérer le plus complètement possible les molécules actives grâce à la salive : glandes parotide (P) submandibulaire (GSM)-autrefois dite sous-maxillaire – et sublinguale (GSL) dont la sécrétion est d’origine réflexe par stimulation du système
parasympathique. (Voir infra) - La sensibilité somesthésique (ou sensitivité) permettant de reconnaitre la texture des aliments qui est portée par la branche mandibulaire du nerf trijumeau ; elle fonctionne comme la sensibilité générale du reste du corps et appartient à une voie dite trigéminolemniscale du fait de sa parenté avec la voie proprement lemniscale.

Les cellules réceptrices se groupent en bouquets appelés bourgeons gustatifs (bg) –fig 3– faits chacun d’une cinquantaine de cellules ; certaines étant gustatives actives caractérisées par la présence de poils gustatifs dirigés vers la cavité bucco-pharyngée ; d’autres plus latérales ne sont pas pileuses et servent à la fois de cellules de soutien et de remplacement aux cellules centrales ces dernières ayant une durée de vie d’une dizaine de jours : l’épithélium stratifié adjacent (esa) fournissant alors les remplaçantes des cellules du bourgeon devenues non fonctionnelles ; à ce titre ce processus de remplacement est très voisin de celui des cellules olfactives. A la base de chaque cellule se détache un axone (a) qui rejoint en profondeur les nerfs gustatifs qui conduiront l’influx nerveux jusqu’au tronc cérébral. Les bourgeons gustatifs des papilles caliciformes du V lingual se situent dans les profonds sillons (s) qui les bordent. –fig 4–


Les voies gustatives sont croisées (fig 5) le côté droit étant traité par l’hémisphère gauche et inversement. Les influx proprement gustatifs sont amenés au tronc cérébral par 3 nerfs crâniens :
- ceux provenant des deux tiers antérieurs de la langue sont conduits par le nerf facial (VII). Sur la fig 5 la branche sensorielle du VII est accolée sur une partie de son trajet à une branche sensitive du V mandibulaire (nm) dont il sera fait une mention plus détaillée en infra ; l’ensemble prend le nom de nerf lingual (nl).
- ceux du V lingual , du tiers postérieur de la langue et de la bouche par le glossopharyngien (IX)
- ceux très postérieurs (épiglotte et pharynx) par le pneumogastrique (X) ;
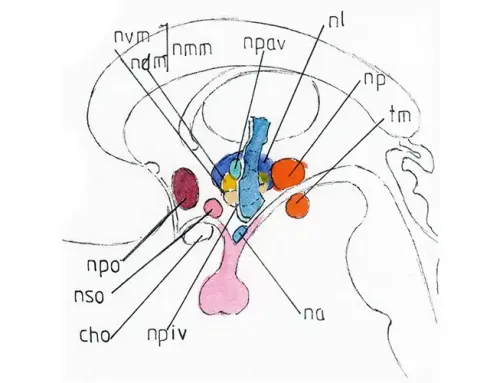
La partie sensorielle de ces nerfs ,représentée ici, se terminent dans un noyau unique et allongé à la partie postérieure du bulbe et de la jonction bulbo-protubérantielle : le noyau solitaire (NS) qui est donc le support intermédiaire de toutes les voies gustatives ; les voies issues du VII se terminant à sa partie haute, celles du X à sa partie basse et celles du IX entre les deux .
Les voies de deuxième ordre entre noyau solitaire et thalamus (Th) croisent la ligne médiane pour se terminer dans un des noyaux ventraux dédiés du thalamus controlatéral ; les voies de troisième ordre empruntent la partie haute de la capsule interne (CI) pour rejoindre le cortex cérébral gustatif situé en profondeur sur la berge pariétale de la scissure de Sylvius (SS) et sur l’insula (I).
La sensibilité générale discriminative (mais non gustative) de la langue et du plancher de la bouche est donc transmise par le nerf lingual (nl) branche du nerf mandibulaire (nm), lui même branche du nerf trijumeau (V ième paire) dont les corps cellulaires se trouvent dans un renflement situé avant sa pénétration dans la protubérance (le ganglion de Gasser GG dit aussi trigéminal) ; nous avons vu que le nerf lingual s’accolait dans sa partie basse avec la branche sensorielle du VII).
Le trajet de cette voie sensitive (et non sensorielle) est marquée par un trait fin ; après connexion avec le très long noyau sensitif du trijumeau (NST) les neurones de deuxième ordre croisent également la ligne médiane pour se rendre au thalamus controlatéral ; de là des neurones de troisième ordre la conduise au cortex dédié à la sensibilité de la langue, de la bouche et du pharynx qui se trouve à la partie la plus basse du cortex somesthésique de la circonvolution pariétale ascendante (CPA).
Le VII véhicule aussi des fibres parasympathiques issues du noyau salivaire supérieur (nsa-s) du bulbe (voir pages sur les nerfs crâniens) et destinées aux glandes sublinguale (GSL) et sub mandibulaire (GSM) : ces fibres végétatives sont représentées en vert sur la Fig 6 ; elles empruntent le même trajet que celui de la branche sensorielle du VII ; elles s’accolent donc elles-aussi au nerf lingual à sa partie basse.


Dans le détail Le VII (fig 6 et 6 agrandie) s’engage dans le méat acoustique interne de l’os du rocher (R), orifice commun avec la sortie du nerf vestibulo-cochléaire (autrefois dit auditif) non représenté sur le schéma ; il chemine à l’intérieur de cet os jusqu’à une formation arrondie appelée ganglion géniculé (gg) et qui représente en fait les corps cellulaires des neurones sensoriels des 2/3 antérieurs de la langue ; ce nom vient du fait qu’à partir de lui le VII se porte brusquement en arrière formant une sorte de genou. Un peu plus bas à hauteur de la caisse du tympan les parties sensorielles et végétatives du nerf se détachent de la partie motrice (en pointillé rouge) pour rejoindre le nerf lingual (nl) issu de la branche mandibulaire (nm) du V ; cette liaison est appelée corde du tympan (ct) en raison de la proximité d’une partie de son trajet avec la face profonde du tympan ; mais elle n’a rien à voir avec lui sur le plan fonctionnel.
Les IX et X sortent de la base du crâne par le foramen jugulaire qu’ils partagent avec la veine jugulaire interne et le XI (non représentés) ; ne sont figurées que les branches sensorielles de ces nerfs, celles du IX étant destinées à la partie postérieure de la langue et celles du X à l’épiglotte et la région pharyngée.
En ce qui concerne l’innervation salivaire, elle vient du VII et du IX :
- celle du VII vient, comme dit plus haut, du noyau salivaire supérieur (nsa-s-), puis suit le trajet du VII jusqu’à la corde du tympan qu’elle emprunte pour rejoindre le nerf lingual ; elle l’abandonnera au niveau du plancher de la bouche pour atteindre le ganglion submandibulaire (gsm) point de relais pour les glandes submandibulaire (GSM) et sublinguale (GSL).
- celle du IX vient du noyau salivaire inférieur (nsa-i-) et transite par ce même nerf qu’elle abandonne dès la sortie du foramen jugulaire pour se porter en avant vers un ganglion également végétatif : le ganglion otique (go) situé au contact du nerf lingual au dessus du raccordement de la corde du tympan ; du go partent vers l’arrière de courtes fibres végétatives jusqu’à la glande parotide (P) la plus volumineuse des 3 glandes salivaires.
On peut ainsi résumer cette innervation du goût de la façon suivante :
- l’innervation sensorielle est assurée par les VII , IX et X ièmes paires crâniennes terminant toutes leurs trajets dans le noyau solitaire (un de chaque côté) avant de relier le cortex
- l’innervation sensitive est assurée par la branche linguale du nerf mandibulaire, lui même branche du V.
- l’innervation salivaire par le VII pour les deux glandes les plus basses (alors que le noyau d’origine est le plus haut), et par le IX pour la parotide, glande la plus haute, son noyau d’origine étant cependant plus bas.