La vision – B – Conduction vers et à travers le cerveau
1 – De la rétine au cortex
La conduction des influx nerveux nés au niveau de la rétine (des 2 rétines !) fait appel à des notions qui ne tombent pas d’emblée sous le sens pour deux raisons ; la principale étant le risque de confondre champ visuel et réception rétinienne, la seconde tenant au caractère en grande partie binoculaire de la vision.
En effet on entend par champ visuel la partie de l’espace que peut capter telle ou telle zone de la rétine quand l’oeil regarde directement devant lui ; les différentes aires réceptrices d’une rétine ont des projections parfaitement logiques mais qui peuvent surprendre à première vue ; en effet chaque rétine peut être assimilée à deux hémisphères vues en creux, jointives par leur équateur vertical et moulée à la face intérieure de l’œil correspondant ; on peut parler ainsi d’hémi-rétines interne et externe.
Une est située du côté du nez (interne, ou mieux nasale ou mieux encore médiale dans le vocabulaire anatomique récent) ; l’autre est dite externe, mieux temporale ou mieux encore latérale ; la délimitation de ces deux hémi-rétines est un équateur vertical passant au point d’émergence du nerf optique ; ainsi la fig 7 montre sur une coupe horizontale des deux yeux les positions médiale (M) et latérale (L) de chaque hemi-rétine.

L’œil gauche, par exemple, ne regarde pas uniquement le champ visuel (CV) gauche (en bleu) mais une partie de celui de droite (en orange foncé) ; pour bien comprendre ce phénomène il faut tirer des lignes de projection à partir du champ visuel et passant par le centre des 2 cristallins et voir où elles se terminent sur chaque hémi-rétine ; la ligne venant de la partie la plus périphérique (point a) du champ visuel gauche (en bleu clair) ne peut se terminer que sur l’hémi-rétine médiale de l’œil gauche, la pénétration dans la pupille de l’œil droit lui étant impossible ; la vision de cet espace ne peut être que monoculaire.
Par contre situé sur un champ gauche plus médian (en bleu plus foncé) le point b peut être vu par les deux yeux, mais sur des hémi-rétines différentes : médiale pour l’œil G et latérale pour l’oeil D : la vision du point b est donc binoculaire ; on peut faire la même observation du côté droit.
Si nous considérons l’hémi-rétine médiale de l’œil G indiquée par un hémi-cercle bleu et l’hémi-rétine temporale de l’œil droit (indiquée aussi en bleu), nous voyons que ces deux hémi-rétines peuvent balayer un champ visuel très important allant du champ gauche le plus latéral jusqu’au champ médian droit dont la limite se trouve entre l’orange foncé et l’orange clair. Idem et inversement pour le côté droit.
Les deux parties foncées du champ visuel (bleu et orange foncés) correspondent à une zone médiale assez étendue qui peut être vue par les deux yeux en même temps : c’est le champ de vision binoculaire (VBO), contrairement aux zones latérales qui sont monoculaires (VMO).
Les choses se compliquent encore un peu si on raisonne en terme de champ visuel inférieur et supérieur (fig 8) ;

en effet plutôt que de scinder la rétine verticalement, on peut la scinder horizontalement et on obtient deux autres hémi-rétines : l’une inférieure, l’autre supérieure ; les projections des points des champs visuels supérieur et inférieur vont se faire sur les hémi-rétines opposées : les deux points h et i de l’hémichamp inférieur se projetteront sur les hémi-rétines supérieures et inversement ; cette inversion haut-bas explique aussi pourquoi les images se forment inversées sur la rétine.

La fig 9 donne montre les 4 couleurs du champ visuel total ; la partie la plus latérale G en bleu clair est vue uniquement par l’oeil G en vision monoculaire ; idem pour l’orange clair et l’œil droit ; l’ensemble des couleurs foncées (bleue et orange) est vu par les deux yeux en vision binoculaire (pour mieux se repérer revenir à la fig 7) ; la forme en cœur du champ binoculaire montre un espace assez nettement plus vaste du champ supérieur par rapport au champ inférieur ; cela vient des projections des parties osseuses du visage notamment du nez qui rogne sur les projections latérales des hémi-rétines supérieures et donc sur les champs visuels binoculaires médians et inférieurs.

La fig 10 va permettre de comprendre le jeu complexe de ces hémi-rétines et de leur projection vers le cerveau.
Avant tout signaler que ce schéma est une simplification anatomique extrême car il est très difficile de suivre correctement le trajet des fibres nerveuses des rétines sans prendre un peu de liberté avec la description anatomique exacte ; il s’agit en effet d’une vue de haut du cerveau alors que les voies optiques se situent à sa face inférieure : il faut donc imaginer un cerveau transparent à travers lequel on voit les voies optiques par le dessus ; de même ont été positionnés en vert clair la position centrale de l’hypophyse (H) et du tronc cérébral (TC) eux-mêmes situés à la face inférieure du cerveau et donc invisibles du haut si ce dernier n’était pas considéré transparent.
Une fois le principe du schéma bien compris il devient possible de décrire le trajet des voies visuelles ; chaque œil donne naissance à un nerf optique (NO) ; ces deux nerfs se rejoignent sur la ligne médiane juste devant l’hypophyse pour former le chiasma optique (CH) ; du chiasma partent les deux tractus optiques (TO) qui contournent le tronc cérébral pour atteindre de chaque côté une formation qui dépend dépend des thalamus : il s’agit des 2 corps genouillés latéraux (CGL) dans lesquels les neurones issus des cellules ganglionnaires de la rétine (voir page A sur la vision) font synapses avec un ensemble de fibres nerveuses de deuxième ordre ; ces dernières appelées radiations optiques (RO), (teintées sur le schéma de différentes nuances de bleu et d’orange) se projettent en arrière vers les 2 aires 17 (aires visuelles primaires) à la partie toute postérieure et interne des lobes occipitaux(voir infra).
Il nous faut alors revenir au couleurs de la fig 7 qui différencient les deux hémi-rétines médiale et latérale ; il a été convenu que l’hémi-rétine médiale de l’oeil G et l’hémi-rétine latérale de l’œil D sont bleues, les symétriques orange ; le phénomène très particulier des voies optiques est que les filets nerveux issus des hémi-rétines latérales ne croisent pas la ligne médiane alors que les médiales la croisent au niveau du chiasma optique ; de ce fait les fibres bleues qui ramènent les influx venant des hémi-rétines bleues participant au même champ visuel gauche passent toutes dans l’hémisphère droit et inversement pour les oranges ; les fibres nerveuses issues de ces deux hémi-rétines complémentaires ont donc un champ visuel controlatéral qui s’arrête sur la ligne médiane ; par conséquent une lésion complète d’un côté des voies visuelles se trouvant en arrière du chiasma entraînera une perte complète du champ visuel du côté opposé et cela jusqu’à la ligne médiane.
Nous avons aussi voulu (encore que très schématiquement) montrer par des couleurs différentes que les fibres formant les tractus optiques transportent des éléments différents suivant l’origine du champ visuel concerné : origine maculaire –fma– (couleur foncée), origines rétiniennes supérieure –frs- (champ visuel inférieur- cf fig 8) origine rétinienne inférieure –fri– (champ visuel supérieur -cf fig 8) : les frs sont les plus hautes ; les fri les plus basses ; les fma se trouvent au milieu ; les fri ont l’originalité de décrire une courbe convexe vers l’avant assez prononcée à la partie postérieure du lobe temporal ce qui peut entraîner une certaine vulnérabilité en cas de lésion pariétale ; les frs et les fri se terminent à la face interne de leur lobe occipital correspondant (les frs au dessus du sillon calcarin -voir infra- ; les fri au dessous) ; les fibres d’origine monoculaire de chaque groupe (couleur très claire) se terminent à la partie la plus profonde (ou antérieure) de la scissure inter-hémisphèrique ; les fibres d’origine binoculaire (plus foncées) en arrière de la précédente à la partie moyenne de la scissure ; les fma (couleur très foncées) se terminent à la partie la plus postérieure de la scissure ainsi qu’un peu au pôle postérieur externe- (voir infra).
Les fig 10 et 11 (lignes rouges) montre l’amputation du champ visuel provoqué par certaines pathologies ; ps : les petites lignes rouges perpendiculaires aux voies visuelles de la fig 10 montrent la situation lésionnelle correspondante à chacun des cas suivants.
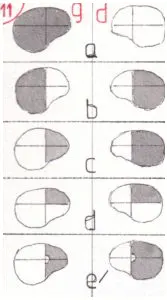 En a la section d’un nerf optique (le G sur le schéma) provoque la perte totale du champ visuel de l’œil gauche mais partiellement compensée par l’intégrité de celle de l’œil droit dont le champ visuel s’étend de la région latérale droite au champ médian gauche compris ; seul le champ latéral G reste invisible. Par contre il n’y a plus de zone en vision binoculaire.
En a la section d’un nerf optique (le G sur le schéma) provoque la perte totale du champ visuel de l’œil gauche mais partiellement compensée par l’intégrité de celle de l’œil droit dont le champ visuel s’étend de la région latérale droite au champ médian gauche compris ; seul le champ latéral G reste invisible. Par contre il n’y a plus de zone en vision binoculaire.
En b l’atteinte du chiasma supprime les deux champs latéraux et conserve la vision binoculaire des champs médians ; des lésions variées peuvent être en cause, la plus classique étant la compression ou l’envahissement du chiasma par une tumeur de l’hypophyse : on parle d’hémianopsie bitemporale.
En c l’atteinte du tractus optique G entraine la perte de vision du champ médian droit de l’œil G et des champs latéral et médian droits de l’oeil D ; or le champ médian droit de l’œil G est le même que le champ médian droit de l’oeil D ; le champ visuel restant n’est donc plus qu’un hémichamp allant de la ligne médiane au champ visuel latéral gauche compris ; la vision reste par contre binoculaire sur le champ médian gauche ; cette atteinte porte le nom d’hémianopsie latérale homonyme : elle se caractérise par la perte complète de l’hémichamp visuel du côté opposé à celui de la lésion du tractus optique considéré ; l’atteinte du champ visuel est donc beaucoup plus importante qu’en cas d’atteinte complète d’un œil. C’est une séquelle qui s’oppose davantage à la possibilité de la conduite automobile qu’une perte complète d’un œil.
En d l’atteinte des radiations optiques donne lieu à des séquelles variées en fonction de leur atteinte complète (donnant la même hémianopsie que précédemment) , ou partielle, ce qui est le cas de certains AVC temporaux qui touchent assez souvent la partie très convexe et antérieure des radiations inférieures (ligne rouge en d) : dans ce cas elle donnera une atteinte des quadrants supérieurs (inversion bas-haut) toujours du côté opposé à la lésion : on parle de quadranopsie.
En e une atteinte complète donnera toujours la même hémianopsie ; mais eu égard à la dispersion des radiations à cet endroit une atteinte partielle est plus fréquente donnant lieu à des séquelles variées ; en e’ on montre une atteinte large mais épargnant les fibres d’origine maculaire provoquant une hémianopsie toujours du même type mais avec une épargne maculaire complète, le côté aveugle ne concernant pas la macula.
2 – Au niveau du cortex visuel
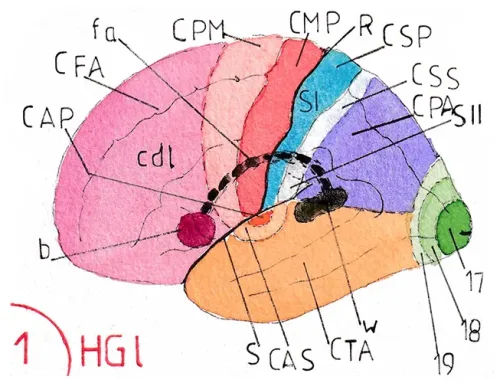
Le lobe occipital est le récepteur des voies visuelles ; les sillons anatomiques qui délimitent les 6 circonvolutions occipitales classiques (de O1 à O6 .cf pages anatomie) n’ont aucune valeur fonctionnelle ; l’ensemble du cortex cérébral a en effet été cartographié en 52 aires d’architectures histologiques différentes par un neurologue allemand du nom de Brodmann en 1909 ; ces aires architecturales se sont avérées correspondre à des aires fonctionnelles beaucoup plus pertinentes que les descriptions anatomiques ; c’est ainsi que les trois aires visuelles correspondent aux aires 17, 18 et 19 de Brodmann.


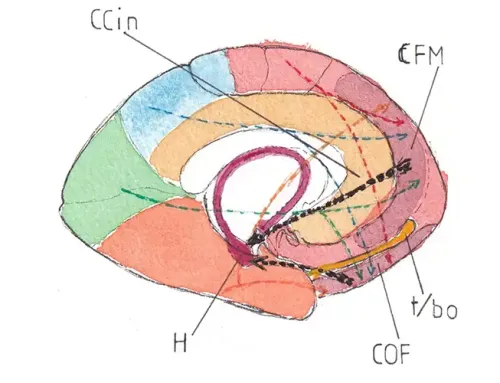
Les fig 12 et 13 montrent la position de ces trois aires sur une face latérale d’un hémisphère (fig12-ici le gauche) et sur une face médiale (fig 13- ici le droit) ; l’aire 17 est l’aire de première réception corticale des influx rétiniens : c’est l‘aire visuelle primaire ; elle est centrée sur un profond sillon visible uniquement sur la face médiale : le sillon (ou la scissure) calcarin (SC) ; les aires 18 et 19 situées concentriquement autour de l’aire 17 sont les aires visuelles secondaires chargées de construire les images intermédiaires ou finales et cela selon des modalités encore encore très mal connues. On peut voir sur ces schémas que l’aire visuelle primaire se trouve pour l’essentiel à la face médiale des lobes occipitaux ; seule une toute petite partie émerge à la pointe extrême de la face latérale de ces lobes.


Les Fig 14 et 15 montrent un agrandissement de la face interne des lobes occipitaux : fig 14 pour le gauche et fig 15 pour le droit ; les 2 aires 17 ont été colorées avec les couleurs correspondantes des fibres des radiations optiques vues précédemment ; ainsi l’aire 17 gauche recevant le champ visuel droit est en orange ; la droite recevant le gauche est en bleu ; la zone d’arrivée des champs monoculaires – VM (donc périphériques) est de couleur claire : elle se situe à la partie la plus antérieure de l’aire, juste en arrière du bord postérieur du corps calleux (CC) ; la zone d’arrivée des fibres d’origine maculaire (M) est de couleur foncée ; c’est la plus postérieure et on peut souligner l’importance de son étendue eu égard à l’extrême petitesse du champ maculaire sur la rétine : cela tient au caractère extrêmement précis de la vision maculaire ; on peut le comparer avec la grandeur de cortex dédié à la main dans les cortex primaires moteur et sensitif. Entre les deux se trouve la terminaison des fibres porteuses des champs médians binoculaires (VB).
En comparant les couleurs des fig 14 et 15 avec celles des radiations optiques de la fig 10 on constate aussi que logiquement les fibres qui conduisent les influx venant des hémi-rétines supérieures (donc des hémichamps visuels inférieurs) se terminent sur la lèvre supérieure du sillon calcarin (SC) et inversement ; donc dans le cortex visuel, comme sur la rétine, l’image est inversée ; cette constatation a pu entrainer quelques interrogations, mais il n’y a ni haut ni bas pour une surface corticale ; aucun renversement n’est nécessaire : seule l’habitude très vite acquise de lier empiriquement le monde matériel et le monde visuel est responsable de cet état de chose ; cette habitude est d’ailleurs modifiable; pour le confirmer il est possible de faire porter à quelqu’un des lunettes inversantes qui ramènent le champ visuel inférieur sur la rétine du bas et le supérieur sur la rétine du haut ; pendant quelque temps le sujet voit effectivement le monde à l’envers ; mais s’il continue à porter ces lunettes quelques jours il le revoit assez vite à l’endroit ; le phénomène inverse se produit quand il les abandonne.

La fig 16 essaie de donner une idée du traitement des messages visuels quand ils arrivent dans le cortex visuel primaire de l’aire 17 ; cette aire présente, comme le reste du cortex, les 6 couches cellulaires mais avec leur structure propre ; les cellules visuelles y sont rangées par colonnes verticales et l’ensemble d’une colonne isolée est sensible à la même sorte et à la même forme de stimulus ; dans la partie gauche du schéma on montre des colonnes sensibles à la direction de traits (—) se trouvant dans le champ visuel à tel ou tel endroit de ce champ ; il peut aussi bien s’agir d’un trait composant une lettre ou d’un bord entre deux surfaces délimitant deux objets différents ; seule sera alors activée la colonne correspondant à la direction du trait en question ; les autres resteront au repos ; sur une toute petite surface se trouvent toutes les colonnes proposant des directions variées et d’ailleurs ordonnées selon une direction angulaire progressivement croissante puis décroissante ; ces « microaires » se reproduisent sur toute la surface du cortex primaire visuel, espacées d’environ 1mm ; la couverture du champ visuel est donc totale ; la couche 4 contient très peu de cellules mais essentiellement des connexions neuronales ce qui laisse à penser que c’est dans cette couche que se fait la jonction entre les axones des fibres des radiations issues des corps géniculés latéraux (cgl) et les dendrites des neurones visuels (nv) ; à partir de cette connexion les influx remontent dans la colonne de neurones jusqu’à émettre des axones qui se connecteront avec d’autres neurones corticaux dans les aires secondaires (as) pour assembler les messages et en faire une image globale ; les étages 5 et 6 semblent dédiés à la transmission des mêmes messages initiaux mais vers des structures sous-corticales (ssc) impliquées notamment dans les mouvements oculaires réflexe.
Dans ce cortex de l’aire primaire ont été aussi découverts d’autres neurones visuels sensibles à d’autres stimulus : notamment au mouvement et à sa direction (symbolisés par des flèches: →), d’autres à la longueur des interfaces (traits de longueurs croissantes et décroissantes: – —) -partie G de la fig 16 – ; d’autres aux couleurs : celles-ci captées par des cônes rétiniens sensibles à différentes longueurs d’onde des rayons lumineux, après passage par les voies visuelles indiquées précédemment, font sans aucun doute relais avec des colonnes sensibles à telle ou telle couleur permettant toute la gamme de mixage et donc de représentation colorée au sein des aires visuelles secondaires. Ainsi l’ensemble du cortex de l’aire 17 est tapissé de colonnes spécialisées couvrant un grand nombre de stimulations primaires venant de la totalité du champ visuel.
La classification de Brodmann est essentiellement une classification histologique mais elle convient imparfaitement aux physiologistes qui lui préfèrent la classification en V ; classiquement de V1 à V5, V1 restant synonyme de l’aire 17 ; certaines dépassent maintenant largement V5 jusqu’à V8 … L’aire V3a a été isolée de l’aire V3 … Tout cela, non pour compliquer les choses mais parce que la recherche est intense dans un domaine qui progresse vite mais est encore loin d’être complètement appréhendé.
Pour rester dans une certaine simplicité on peut dire que l’aire V2 qui correspond à l’aire 18 est certainement celle qui permet de réunir les images primaires détectées par V1 et qui commence à former des images plus interprétables ; mais ce qui se passe après reste du domaine de la recherche .
Une chose parait acquise : c’est l’enchaînement d’aires adjacentes sur un rayon dorsal qui permet la localisation dans l’espace et celui sur un rayon ventral pour la reconnaissance des formes :

Fig 17 : le rayon dorsal partant de V1, passe par la partie dorsale de V2, puis la partie supérieure et latérale de l’aire 19 dénommée V5 pour aboutir sur les aires 5 et 7 de Brodmann qui appartiennent au lobe pariétal et dont on connait l’importance pour la localisation dans l’espace ; c’est le circuit du « où ? » qui permet non seulement de s’orienter visuellement dans l’espace mais aussi de suivre l’évolution des changements qui s’y produisent ;
Des lésions de ce circuit peuvent entrainer des troubles très curieux comme le cas d’une personne qui après avoir fait un AVC sur ce territoire ne voyait plus l’écoulement du thé qu’elle versait dans sa tasse ; le liquide lui apparaissait comme figé ne s’écoulant pas et bien sur elle ne voyait pas non plus sa tasse se remplir et déborder.
Fig 18 : c’est le « circuit du quoi ? » permettant de reconnaitre notamment les objets et les visages ;
Il passe par la partie inférieure et médiale de V2, celle de l’aire 19 (ou V4) et se prolonge sur la face inférieure du lobe temporal notamment dans la circonvolution occipito- temporale médiane appelé gyrus fusiforme –gf–( V4 +aire 37 ) ; V4 dans sa partie inférieure et médiale semble capitale pour percevoir les couleurs et les objets ; l‘aire 37 pour la découverte des visages ; les aires 20 et 38, proches de l’amygdale (A) pourrait faire le lien entre peur et visages inhabituels ou menaçants. Les aires 37, 20 et 38 appartiennent au lobe temporal. Ces deux trajets complémentaires montrent bien qu’une vision complète et précise déborde le lobe occipital et nécessite l ‘activation étendue de certaines aires pariétales et temporales.