La vision – A – Partie optique
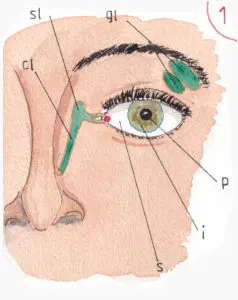
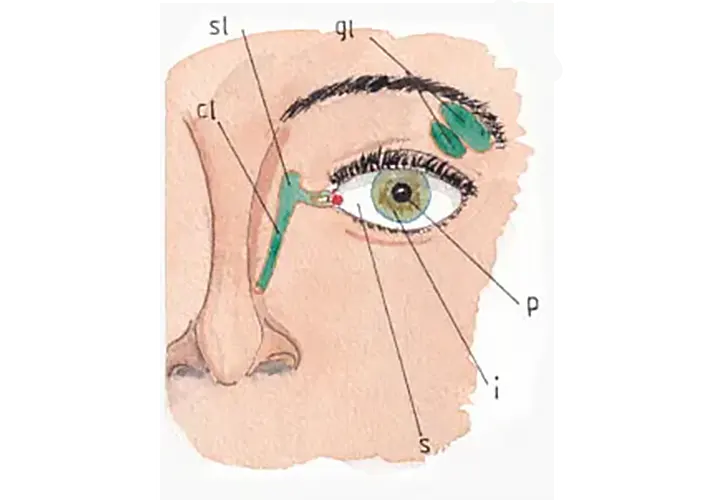
La fig 1 permet de revoir les notions anatomiques de base : la sclérotique (s) en blanc constitue l’enveloppe extérieure de l’œil ; l’iris (i) est la couronne colorée recouverte par la cornée non visible sur le schéma car totalement transparente ; l’iris délimite un petite surface noire parfaitement circulaire : la pupille (p) par où pénètrent les rayons lumineux ; la taille de la pupille est en permanence réglée par l’ouverture du cercle le plus interne de l’iris (voir infra).
Notons sur cette figure la projection des glandes lacrymales (gl) avec leurs portions sous orbitaire et palpébrale qui sécrètent les larmes et les déversent par de micro-canalicules à la partie profonde de la paupière supérieure ; elles sont sécrétées en continu pour assurer la lubrification de la face externe de l’oeil dans ses mouvements à l’arrière des paupières ; cette sécrétion dépend du nerf facial grâce à une branche nerveuse très fine issue de son trajet végétatif parasympathique et dont le point de départ est le noyau salivaire supérieur (v page sur les nerfs crâniens).
Les larmes sont récupérées à l’angle interne de l’œil par deux petits canaux qui les conduisent au sac et au conduit lacrymal (sl et cl) avec une évacuation dans la fosse nasale correspondante.
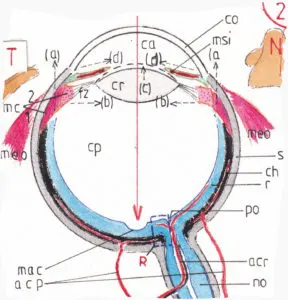
NB : les épaisseurs des différentes tuniques montrées sur ce schéma ne correspondent pas à la réalité anatomique.
La fig 2 montre une coupe horizontale du globe oculaire gauche : bien qu’à une échelle plus petite N indique la place du nez et T celle de la tempe gauche. On retrouve l’enveloppe extérieure de l’oeil, la sclérotique (s) en grisé sur le schéma ; elle bombe en avant tout en devenant transparente pour former la cornée (co) ; l’enveloppe moyenne est la choroïde (ch) très riche en mélanine pigment noir qui permet à la rétine (r) (enveloppe interne) de fonctionner comme si elle était à l’intérieur d’une chambre noire donc à l’abri de toute diffraction des rayons lumineux.
Les rayons lumineux (flèche rouge →) doivent traverser 4 couches de densité différente avant d’atteindre la rétine : la cornée, la chambre antérieure (ca) milieu liquide, le cristallin (cr) et la chambre postérieure (cp) remplie d’un gel épais l’humeur vitrée. Chacune de ces couches réfracte la lumière en fonction de l’angle d’attaque du rayon lumineux, de la densité propre de chaque tunique et plus encore de la différence de densité entre deux couches adjacentes ; à noter que seuls les rayons lumineux centraux (comme celui représenté par la flèche rouge) ne sont pas réfractés car perpendiculaires aux tuniques rencontrées, mais seulement ralentis ; plus ils s’éloignent de ce centre plus la réfraction est importante ; c’est en fait la cornée qui, bien qu’un peu moins dense que le cristallin, réfracte le plus car elle se situe juste après un milieu très peu dense qui est l’air ; le cristallin plus dense réfracte moins car il se situe après un milieu liquide relativement dense ; cependant il a une fonction adaptatrice essentielle : il peut modifier sa courbure notamment antérieure et plus celle-ci est marquée plus il réfracte les rayons lumineux.
Quel en est le mécanisme ?
Le cristallin a une structure lamellaire qui lui permet de changer de forme en fonction de la traction qui est appliquée sur son bord périphérique ; laissé à lui-même son élasticité le rétracte sur lui-même lui donnant une forme plus arrondie ; cette tension est sous la dépendance d’un muscle intrinsèque : le muscle ciliaire (mc) lui même formée de deux parties : la partie méridienne (1) et la partie circulaire (2) ; paradoxalement c’est quand ses deux parties sont relâchées qu’elles elles tirent le plus sur des fibres zonulaires (fz) qui sont solidaires du bord externe du cristallin ; dans ces conditions celui-ci est étiré et ses surfaces en partie aplanies ; à l’inverse quand elles se contractent la partie méridienne attirent les fibres zonulaires en avant (↑a) et la partie circulaire se rapproche du bord cristallinien (→b) ; le cristallin prend alors une forme plus arrondie (↑c – – -) et devient donc plus réfractif ; cette mécanique à priori contre-intuitive vient du caractère circulaire de ce muscle dont la contraction centralise davantage son bord interne ; ce phénomène appelé accommodation est à la base de la vision de près ; il est souvent complété par une diminution du diamètre de la pupille par contraction du muscle sphincter de l’iris-msi– (→d←) qui limite l’entrée des rayons lumineux ; en effet dans la vision de près ils risqueraient de ne pas se concentrer suffisamment vers les points précis concernés de la rétine (voir infra) ; cette double action est en partie assurée par le noyau végétatif de la III ième paire crânienne (nerf oculomoteur) ; elle est donc automatique non soumise à la volonté. La contraction pupillaire à une lumière forte, ou reflexe photomoteur est un phénomène différent entièrement assuré par ce noyau végétatif.
Le schéma montre que la rétine subit une dépression cupuliforme à sa partie centrale : c’est la macula (mac) qui est le point où la vision est la plus précise ; au centre de cette cupule se trouve une zone de réception encore plus précise : la fovéa indispensable a une vision discriminative parfaite ; les deux parties latérales de la rétine (l’une appelée champ nasal, l’autre champ temporal) se rejoignent en un point où les axones des neurones rétiniens se recourbent vers le pôle postérieur de l’œil pour former la partie initiale du nerf optique (no) ; ce point un peu décalé en dedans de l’axe médian est la papille optique (po) ou tâche aveugle car elle n’a plus de cellules réceptrices en regard.
La vascularisation interne de l’œil est assurée par deux systèmes : celui de l’artère centrale de la rétine (acr) pénétrant directement dans le nerf optique et s’épanouissant sur la papille optique pour vasculariser directement la rétine ; celui des artères ciliaires postérieures (acp) perforant la sclérotique pour vasculariser la choroïde : ces deux systèmes, issus d’une même branche l’artère ophtalmique elle-même issue de l’artère cérébrale moyenne ont une importance partagée dans la vascularisation de la rétine.
Deux points notables sur la fig 2 : la présence de l’insertion de deux muscles extrinsèques de l’œil (meo) –voir page vision C – et d’un petit repère rectangulaire en pointillé (R) à proximité de la papille optique et qui servira à l’étude de la structure histologique de la rétine (voir infra).
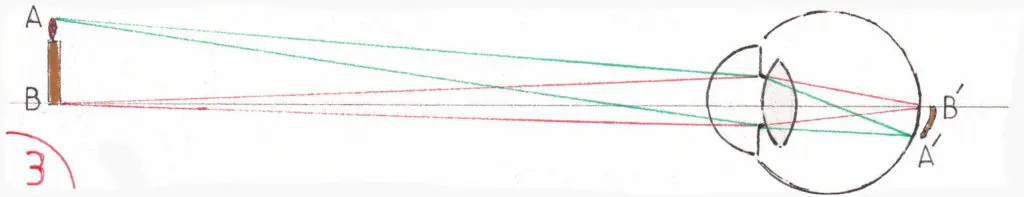
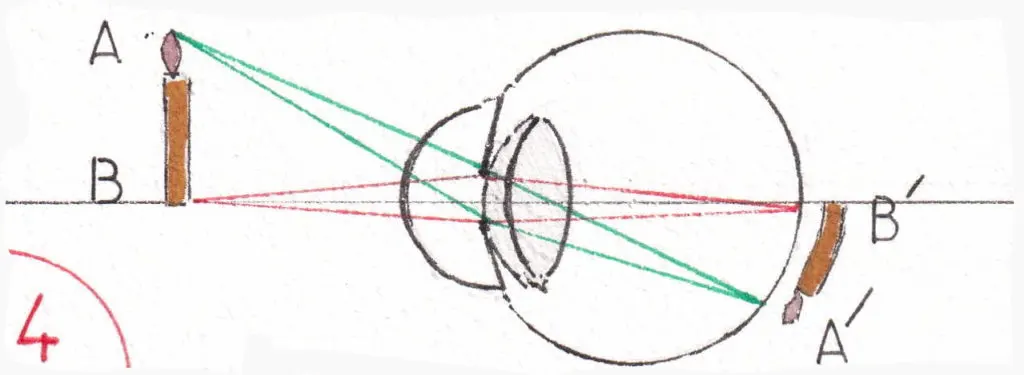
Les fig 3 et 4 montrent le trajet des rayons lumineux issus d’un objet quelconque (ici une bougie) en fonction de l’éloignement de l’objet ; précisons d’emblée que tous les schémas illustrant ce trajet sont simplifiés en ne détaillant pas les réflexions des différents milieux oculaires traversés signalés plus haut ; on fait comme si seul le cristallin en était responsable en donnant à l’ensemble un indice de réfraction de 59 dioptries sans accommodation et correspondant à la somme algébrique de la puissance réfractive de chaque milieu.
La fig 3 montre un objet éloigné ; le cristallin est au repos (en réalité tendu par les fibres du muscle ciliaire) et la pupille largement ouverte ; la construction géométrique explique la convergence des rayons partant des limites extrêmes de l’objet (A et B) en A’ et B’ sur la rétine : l’image rétinienne de l’objet est donc inversée et le restera jusqu’au cortex cérébral récepteur.
La fig 4 montre le même objet rapproché ; le cristallin bombe du fait de la contraction du muscle ciliaire (cf supra) et la pupille se rétrécit limitant l’entrée des rayons ce qui affine leur arrivée sur la rétine et évite une projection imprécise sur elle en cas de réfraction et de sphéricité imparfaites du globe oculaire ; l’image est plus grande (expliquant le grossissement des images quand un objet se rapproche) mais reste toujours inversée.
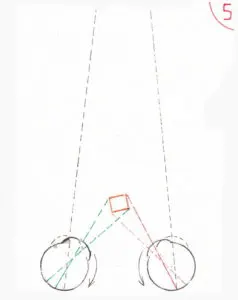
La fig 5 donne une explication (partielle) de la notion de perception de relief favorisée par la vision binoculaire ; en fait une vision monoculaire ne fait pas disparaître la perception de relief qui se base sur un nombre important d’acquis mémorisés : la taille plus petite des objets éloignés mais connus, l’arrêt des rayons lumineux par un objet proche masquant en partie un objet plus éloigné suffisent à donner cette perception ; mais il est incontestable que la vision binoculaire améliore considérablement cette sensation et d’autant plus que l’objet se rapproche ; en effet dans ces conditions la convergence progressive des globes oculaires fait que chaque œil voit une partie un peu différente de l’objet ; trop proche de l’œil l’objet est vu dédoublé car la différence de vue est trop importante pour être interprétée correctement par le cerveau ; mais à une distance convenable le cerveau parvient à établir une différence acceptable et cohérente qui améliore nettement la sensation de relief.
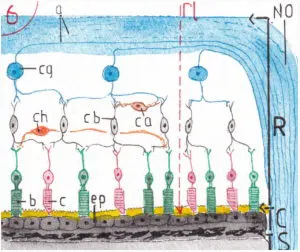
La fig 6 correspond à la coupe histologique centrée sur la petite zone rectangulaire R de la fig 2 ; il s’agit pour l’essentiel d’une coupe histologique de la rétine (R) mais aussi de la choroïde (C) et d’une partie de la sclèrotique (S) ; l’endroit choisi est celui où les axones des cellules portant les messages visuels se regroupent pour former le début du nerf optique (NO) ; l’intérieur de l’œil est donc orienté vers le haut du schéma.
Les cellules réceptrices des rayons lumineux (bâtonnets –b– en vert et cônes –c– en rouge) sont donc les cellules les plus externes ; elles reposent sur les cellules de l’épithélium pigmentaire (ep) de la choroïde jouant le rôle de chambre noire par leur richesse en mélanine ; cette disposition peut paraître curieuse car obligent les rayons lumineux à passer à travers trois couches axono-cellulaires pour atteindre cônes et batonnets : celle des axones (a) du futur nerf optique (en bleu) ; celle des cellules ganglionnaires (cg) dont sont d’ailleurs issus ces axones et celle des cellules bipolaires (cb) faisant le lien entre cônes et bâtonnets d’une part et cellules ganglionnaires d’autre part ; à la jonction des cellules réceptrices et des cellules bipolaires se trouvent des cellules transversales dites cellules horizontales (ch en orange) et à la jonction entre cellules bipolaires et cellules ganglionnaires les cellules amacrines (ca) dont le rôle est encore mal compris.
PS : Le rayon lumineux (rl) présenté en pointillé rouge semble éviter toutes les couches ; ce n’est bien sur pas le cas dans la réalité les cellules étant toutes contigües serrées les unes contre les autres.
Les cônes et les bâtonnets sont donc les cellules réceptrices des rayons lumineux et cela grâce à un composé photosensible associant une protéine fabriquée par la cellule elle-même et une molécule issue de la vitamine A, le rétinal, la choroïde jouant le rôle de réservoir de cette vitamine ; ce composé photosensible se différencie par sa composante protéique suivant que l’on a affaire à un bâtonnet où à l’un des trois types de cônes chacun des trois étant sensible à une couleur particulière (ou plus exactement à la longueur d’onde du rayon incident).
Le principe général de fonctionnement est le suivant : le composé photosensible est porté par les replis membranaires de la cellule (petites stries visibles sur le schéma) situés sur sa partie la plus externe ; l’arrivée d’un rayon lumineux en frappant cette région provoque une modification du rétinal qui se détache de sa protéine porteuse en provoquant des réactions ioniques complexes dans le corps cellulaire du bâtonnet ou du cône concerné ; réactions qui se répercuteront dans les cellules bipolaires puis dans les cellules ganglionnaires et leurs axones, donc in fine dans le cortex cérébral.
Les bâtonnets et les cônes ont une répartition très inégales suivant les régions rétiniennes ; leur rôle est également très différent ; les bâtonnets ne fonctionnent qu’en éclairage de très basse intensité car ils sont inhibés par la lumière qui empêche leur chargement en photo-pigments ; celui-ci ne se fait complètement que dans l’obscurité expliquant le délai d’attente nécessaire pour commencer à discerner la nuit après avoir éteint la lumière ; ils sont donc seuls responsables de la vision nocturne ; ils sont presque exclusifs dans les régions périphériques de la rétine ; ils ne perçoivent pas les couleurs.
Les cônes au contraire ne fonctionnent que que sous un éclairage suffisant ; leur réception très fine leur permet de réagir de façon très précise aux formes et aux couleurs ; ils sont très résistants à la lumière reformant en permanence le couple protéine-rétinal leur permettant une fonction quasi continue ; leur capacité à reconnaître les couleurs vient de la spécialisation préférentielle de chacun des trois types de cônes ; on parle des cônes rouges, bleus et verts ; il vaudrait mieux parler de leur sensibilité préférentielle aux différentes longueur d’onde des rayons lumineux (déterminantes dans la formation des couleurs) qui quand elles se chevauchent donnent toute l’étendue du spectre coloré interprétable par le cerveau ; on est cependant loin d’avoir parfaitement compris l’intégration de toutes ces fonctions permettant l’identification de telle ou telle couleur ; les cônes sont largement préférentiels au centre de la rétine notamment au niveau de la macula et plus spécialement encore de la fovéa ; à ce niveau il n’y d’ailleurs plus que des cônes très petits et très nombreux, chacun étant alors relié séparément à une cellule bipolaire et à une cellule ganglionnaire pour éviter toute déperdition d’information ; si la région de la macula parait déprimée c’est justement par la raréfaction des couches sus-jacentes qui se trouvent déportées latéralement mais restent reliées aux cônes par de longs prolongements axono -dendritiques ; de plus dégagés de ces couches internes les rayons lumineux atteignent les cônes maculaires presque directement.
Pour bien voir il faut aussi bien apprécier les limites des structures variées que nous regardons ; les physiologistes s’intéressent beaucoup à ce qui permet de bien discerner les contrastes ; ce n’est ni la couleur ni la forme d’un objet qui nous permet d’apprécier ses limites, mais ses contrastes par rapport aux structures voisines ; par exemple voir un film en noir et blanc est très suffisant pour comprendre parfaitement une scène visuelle ; des découvertes très importantes ont été faites dans ce sens sur les cellules ganglionnaires, qui le plus souvent recueillent les influx de nombres de cellules photoréceptrices ; elles paraissent fonctionner selon deux modes : un mode « on » (ouvert) et un mode « off » (fermé) et non selon un mode intermédiaire ; on peut trouver là une explication élégante à la façon dont la rétine permet au système visuel d’obtenir des contrastes nets ; en effet si une cellule ganglionnaire dite « on » reçoit des influx lumineux venant du centre de son champ récepteur (déterminé par le nombre de cellules réceptrices qui lui sont reliées) elle répondra par un message excitateur vers son axone, message excitateur qui sera donc conduit au cerveau ; par contre si l’influx lumineux dépasse le centre du champ ou ne l’atteint que latéralement elle restera silencieuse ; selon le même principe une cellulaire ganglionnaire à centre « off » ne réagira que si le centre de son champ est nettement sombre et non si le champ l’est totalement ou sur un côté ; ainsi les deux types de cellules ganglionnaires étant répartis harmonieusement (bien que les off soient les plus nombreuses) il ne peut pas y avoir de transmission vers le cerveau de limites lumineuses ou foncées (les plus nombreuses dans la nature) vagues ou imprécises.
Il semble que les cellules horizontales qui sont toujours inhibitrices mais dont les grandes longueurs dendritiques peuvent réunir des cellules photoréceptrices éloignées puissent jouer également un rôle dans ces processus de contraste très importants pour distinguer avec rapidité les éléments essentiels d’une scène visuelle quelconque.