Encéphale et motricité • 4 • Le cervelet
Le cervelet constitue la troisième structure de l’encéphale , après les noyaux gris centraux et les voies extra-pyramidales à accompagner la pleine réalisation du mouvement volontaire , en participant à l’équilibre du corps (archéo-cérébellum) , en contrôlant le tonus musculaire des muscles engagés dans l’activité posturale (paléo-cerebellum) et en coordonnant dans le temps et dans l’espace les composantes de ce mouvement pour lui assurer le maximum de précision (néo-cérebellum).
Les mots « savants » utilisés pour distinguer ces trois parties du cervelet font une part importante à la phylogénèse , laissant supposer que le cervelet humain s’est construit par strates successives ; le premier faisant son apparition chez le poisson pour qui l’équilibre en milieu liquide est le plus essentiel et le troisième étant particulièrement développé chez l’homme car adapté à la réalisation des mouvements les plus complexes.
Ces trois parties ne recouvrent pas totalement l’anatomie détaillée dans les 2 pages qui lui sont consacrées au chapitre anatomie, même si les comparaisons sont intéressantes :
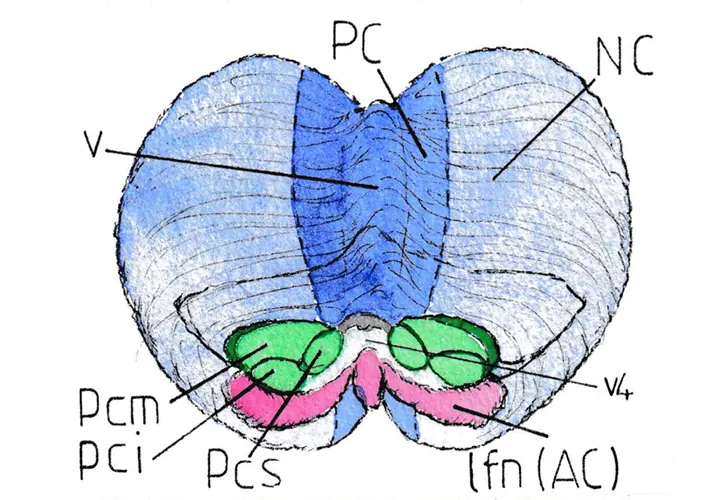
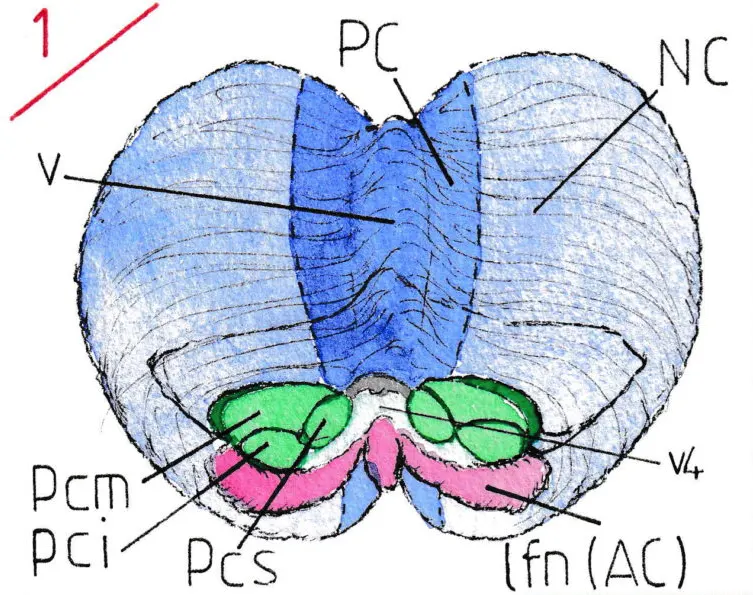
La figure 1 est une vue légèrement distordue par rapport à la réalité pour pouvoir montrer conjointement la face antérieure du cervelet normalement fixée au tronc cérébral par ses 6 pédoncules cérébelleux (ici sectionnés) ainsi que la majeure partie de sa face supérieure. Les pédoncules cérébelleux apparaissent en vert (moyens , supérieurs et inférieurs : pcm , pcs et pci ) encadrant le quatrième ventricule (V4). Au dessous (en violet) se trouve inchangé par rapport à l’anatomie de base, le lobe floculo-nodulaire (lfn) qui constitue l’archéo-cervelet (AC) ; sur la face supérieure le vermis -v- (sorte de sur-élèvement médian) et ses parties adjacentes correspondent au paléo-cervelet (PC) en bleu foncé ; en bleu clair sont représentées les deux larges hémisphères cérébelleux du néo-cervelet( NC). Ces deux aires d’activité différente se poursuivent sur la face inférieure comme le schéma le laisse supposer.
La figure 1a montre en extrêmement simplifié la structure en réalité très complexe du cortex cérébelleux ;
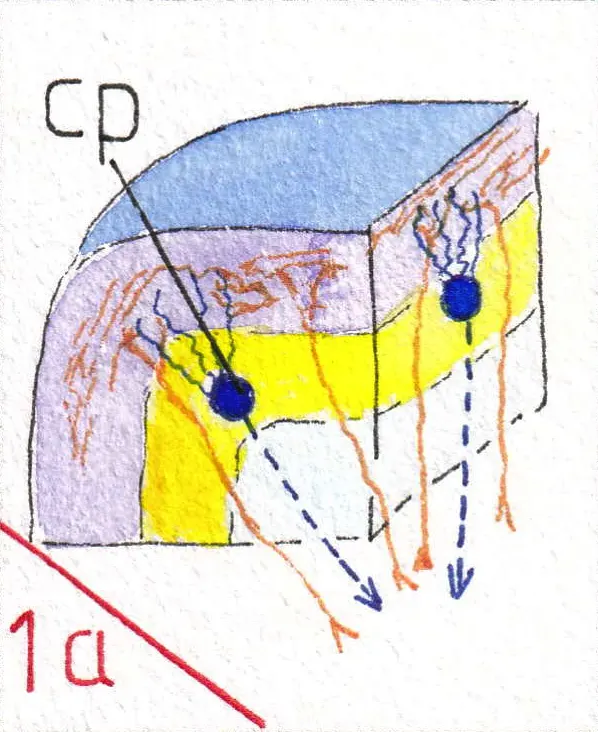
ici n’a été placée que la cellule qui le caractérise à la fois morphologiquement et fonctionnellement : la grosse cellule de Purkinje (cp) (en bleu) qui reçoit les afférences d’une multitude de sources du système nerveux central (en orange) moelle épinière comprise, et dont le rôle (fig 1b) est d’envoyer des influx inhibiteurs (-) plus ou moins importants vers les noyaux cérébelleux (nc) qui sont les seules voies de sortie du cervelet après réception de ces signaux.
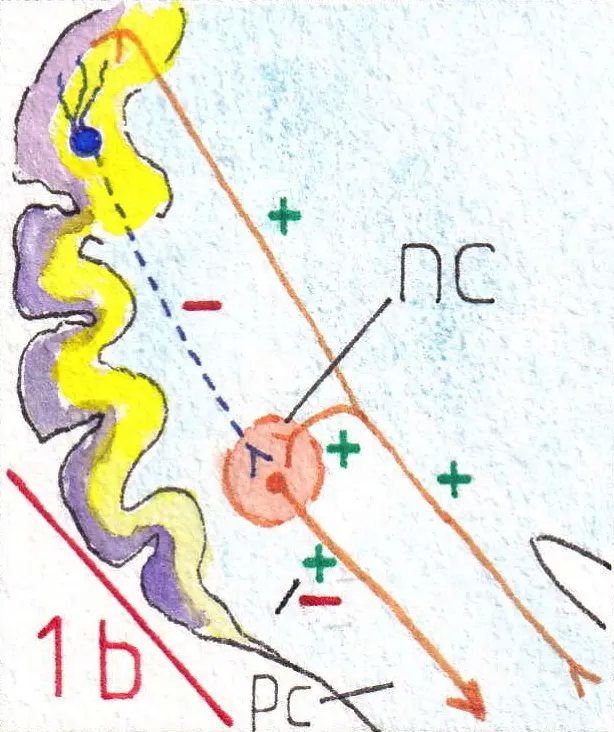
Cependant l’organisation générale montre (fig 1 b) que ces influx sortants sont en général excitateurs car lorsqu’une fibre afférente transitant par un pédoncule cérébelleux (pc) entre dans le cervelet , elle envoie une collatérale excitatrice (+) vers le noyau concerné avant d’atteindre le cortex ; lequel par les cellules de Purkinje lui enverra une efférence inhibitrice (-); la réponse efférente du noyau est donc excitatrice mais contrôlée par le cortex cérébelleux, donnant une réponse modulée ( + – ).
Une caractéristique du cervelet est que l’ensemble du cortex fonctionne sur le même schéma (à l’inverse du cortex cérébral) bien que les noyaux de sortie soient différents pour chacune des trois voies .
1) Les voies de l’archéo-cervelet (fig 2 -profil- et 2a-face-)
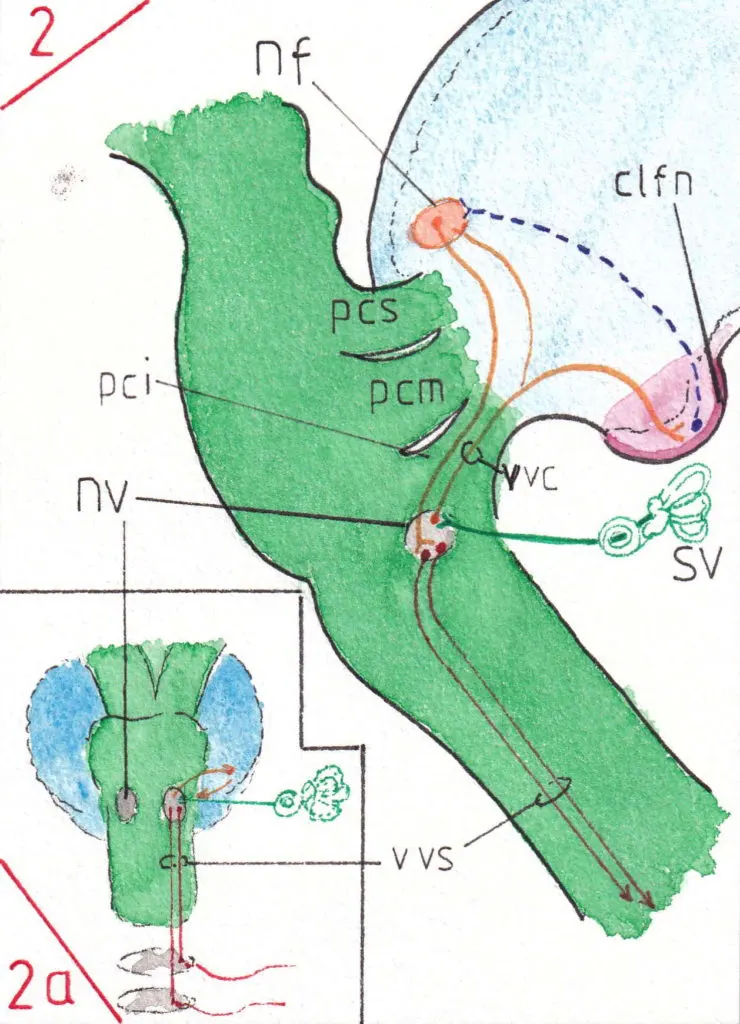
Elles complètent l’activité des voies extra-pyramidales vestibulo-spinales (voir la page voies extra-pyramidales); le noyau vestibulaire (nv) reçoit en effet ses afférences du système vestibulaire (sv) situé dans l’oreille interne ainsi que celles de la sensibilité proprioceptive des muscles , tendons et articulations du corps (voir la page 3 de la motricité ); la voie vestibulo -cérébelleuse (vvc) passe par le pédoncule cérébelleux inférieur (pci) et atteint le noyau fastigial (nf) (qui lui est dédié) ainsi que le cortex du lobe floculo-nodulaire -clfn- (ici le nodule vu en coupe) ; lequel transmet des influx inhibiteurs (bleu pointillé) à ce même noyau (par les cellules de Purkinje) avec une réponse adaptée (par le même pédicule) vers le noyau vestibulaire de départ ; celui-ci se charge de l’activité des muscles axiaux et des ceintures par ses voies vestibulo-spinales (vvs déjà vues page 3 de la motricité); Ainsi cette voie contribue -t-elle à renforcer la fonction d’équilibre et de tonus musculaire du même côté du corps (homolatéral).
2) Les voies du paléo-cervelet (fig 3 -profil- et 3a-face-)
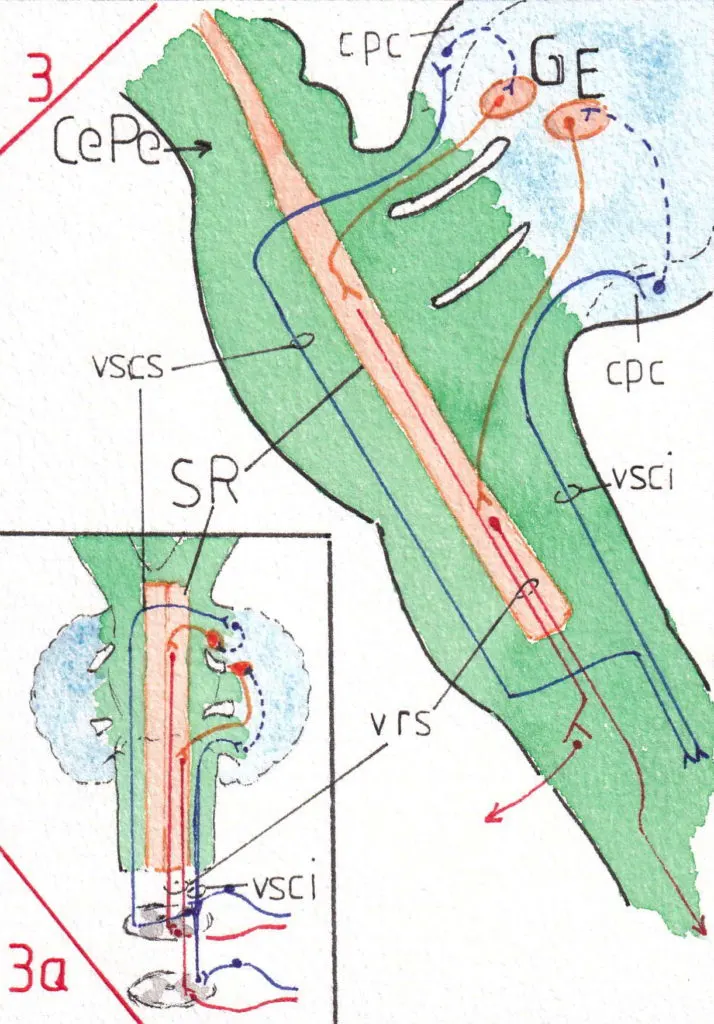
Elles contribuent à l’activité des voies réticulo-spinales (voir la page 3 motricité ) et notamment à la régulation du tonus des muscles engagés dans une action automatisée ; nous avons vu que la voie réticulaire seule était en grande partie dépendante des influx de la voie de la motricité volontaire cortico-spinale notamment du cortex prémoteur ; avec les voies spino-cérebelleuses (vsci et vscs) une précision supplémentaire venant de la périphérie est prise en compte; en effet ces voies extrêmement rapides transmettent au cervelet les influx proprioceptifs inconscients en provenance des muscles tendons et articulations de l’hémicorps homolatéral (même côté) ; la voie en provenance de la partie inférieure du corps (vsci) reste homolatérale sur tout son trajet et s’engage dans le pédoncule cérébelleux inférieur ; celle provenant de la partie supérieure (vscs) du corps croise la ligne médiane dans la moelle mais la recroise à nouveau à la hauteur du pédoncule cérébral (Pe Ce) pour pénétrer dans le cervelet par le pédoncule cérébelleux supérieur (pcs) .
Après relais dans les noyaux cérébelleux correspondants (globulus-G- et embolus -E-) et dans le cortex du paléo-cervelet (cpc) , les voies de sortie vers la substance réticulée (SR) se font par leur pédoncule cérébelleux d’arrivée.
Ainsi le cervelet participe t-il à augmenter le caractère excitateur des voies réticulo-spinales (vrs) ainsi que leur précision qui se trouve encadrée à la fois par le cortex prémoteur et la sensibilité périphérique inconsciente.
Là encore l’action est homolatérale
3) Les voies du néo-cervelet (fig 4 -face- et 4a -coupes horizontales)
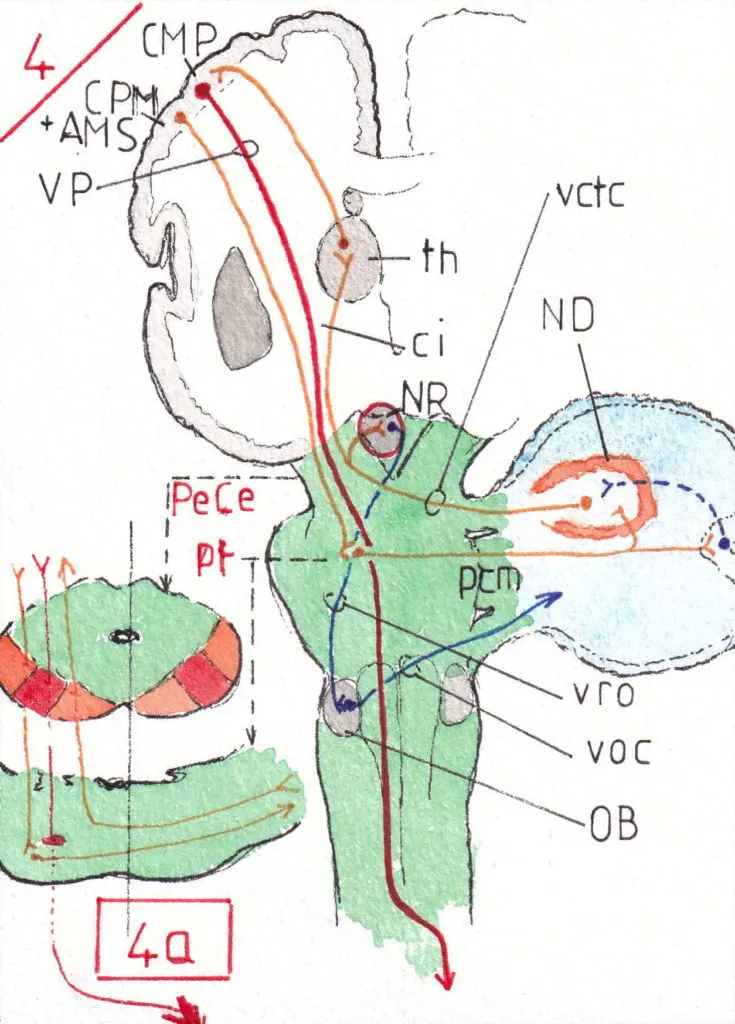
Elles gèrent la précision et la rapidité de nos mouvements les plus soumis à la motricité volontaire notamment la plus périphérique .(membres supérieurs , mains … mais aussi langage articulé!)
Le point de départ se situe essentiellement dans le cortex prémoteur (CPM) et l‘aire motrice supplémentaire (AMS) dont on a vu que la mise en route précédait souvent celle de la voie pyramidale volontaire (VP) ; de là ces fibres dites cortico-pontiques (en orange) accompagnent cette dernière suivant le même trajet : cad capsule interne (ci) et pédoncule cérébral (PeCe) dans lequel elles circulent en dedans et en dehors de la voie pyramidale (fig4a) ; le nombre des axones qui les constituent est infiniment plus grand que pour cette dernière : c’est dire leur importance.
Arrivés au niveau du pont (pt) elles font relais dans une multitude de petits noyaux homolatéraux puis traversent horizontalement la ligne médiane pour s’engouffrer dans le pédoncule cérébelleux moyen (pcm) du côté opposé ; dans cette traversée du pont elles ont dissocié le faisceau pyramidal (VP) en multiples faisceaux qui ne se réunifieront que dans le bulbe .
Dans le cervelet elles se projettent sur leurs noyaux (noyaux dentelés ND) et sur le cortex du néo -cervelet selon le même principe général; la voie de sortie passe alors par le pédoncule cérébelleux supérieur vers le cortex moteur primaire (CMP) du côté opposé avec un relais thalamique (th) ; ce retour permet de contrôler l’action de la voie pyramidale en l’adaptant exactement à ce qu’avait prévu le cortex prémoteur ; on dit que le néo -cervelet « ajuste le mouvement volontaire en comparant le projet moteur à sa réalisation effective «.
Dans cette activité de contrôle de précision la réponse se fait encore dans l’hémicorps homolatéral à l’hémisphère cérébelleux atteint ; en effet l’hémisphère droit ici représenté mobilise l’hémicorps gauche ; mais si l’hémi-cervelet gauche est atteint c’est donc le même côté du corps (ici le gauche) qui sera perturbé . Dans les trois voies étudiées on peut constater que les perturbations d’un hémicorps par atteinte du cervelet sont toujours homolatérales à la lésion contrairement aux lésions cérébrales qui touchent toujours l’hémicorps controlatéral.
Il est important aussi de signaler les efférences de la voie néo-cérébelleuse vers les noyaux rouges (NR) situés à la partie haute des pédoncules cérébraux (PeCe); en effet les noyaux rouges sont connectés avec le retour des voies cérébello-thalamo-corticales(vctc) et une importante voie rubro – olivaire (vro) reliant le noyau rouge à l’olive bulbaire (OB) est probablement à l’origine d’un apprentissage moteur facilitant les mêmes mouvements ultérieurs; en effet l’olive bulbaire est un centre d’afférence venant de la presque totalité du système nerveux central et les liaisons olivo-cérébelleuses (voc) , passant par le pédoncule cérébelleux inférieur, pourraient maintenir en mémoire de nombreux mouvements devenus alors « prêts à l’emploi ».
Ces quelques explications peuvent aider à comprendre les troubles cliniques relevant de lésions du cervelet : l’atteinte globale du cervelet comme cela est vu dans certains TC occipitaux peut entrainer une hypotonie musculaire diffuse très difficile à corriger ; est en cause l’altération diffuse des noyaux cérébelleux de sortie dont on a vu la rôle très important dans le renfort de la tonicité musculaire.
L’atteinte préférentielle des voie néo-cérébelleuses perturbent fortement les mouvements de précision et les mouvements rapides ; elle est responsable de :
- l’hypermétrie : le mouvement dépasse son but; l‘ataxie cérébelleuse : le mouvement volontaire est complètement incoordonné
- l’adiadococinésie : altération des mouvements rapides alternatifs (battement des mains par exemple) qui en raison de démarrages incontrôlés (trop tôt ou trop tard) ne peuvent plus être synchronisés.
- Dysarthrie : atteinte de la coordination de tous les muscles concernés par le langage entrainant une élocution tantôt trop faible , tantôt trop forte explosive pouvant aller jusqu’à une élocution complètement incompréhensible
- Tremblement intentionnel qui s’aggrave à mesure que le sujet s’approche d’une prise manuelle en raison de l’altération de l’amortissement permis par l’intégrité du néo-cervelet
- Troubles de l’équilibre et nystagmus oculaire (tremblement oculaire surtout dans le regard latéral) dans les lésions de l’archéo-cervelet
- Phénomènes de rebond quand on demande au sujet de forcer sur un geste contre résistance ; lors de la levée brusque de cette dernière le geste poursuit son impulsion très au delà de la normale.