Encéphale et motricité • 3 • les voies extrapyramidales
Le terme « voies extrapyramidales » désigne l’ensemble des structures indispensables à une motricité efficace et harmonieuse en appoint de la voie pyramidale volontaire ; sans elles ou en cas de lésion cette dernière est lourdement entravée. Les voies extrapyramidales gérant une automaticité motrice allant des mouvements les plus fins (cervelet) aux plus globaux ; en ce sens les noyaux gris centraux font déjà partie de ces voies ; il en va de même du cervelet étudié plus loin ; mais par habitude le terme de voies extrapyramidales est en général réservé aux fibres motrices qui parcourent le tronc cérébral et relient les hémisphères à la moelle épinière , comme le fait la voie pyramidale , mais d’une façon non ou peu soumise à la volonté.
La description complète des voies pyramidales est impossible dans un court texte de simplification ; certaines de ces voies paraissant par ailleurs avoir un usage redondant et d’autres devenues sans doute plus ou moins utiles alors qu’elles représentent chez nombre d’animaux des fonctions essentielles; notamment chez les quadrupèdes assurant l’automatisme des 4 membres dans la marche et la course.
Nous nous limiterons seulement à deux voies principales : la voie vestibulaire et la voie réticulaire ; la première est surtout centrée sur le maintien de l’équilibre en position debout ; la seconde sur la préparation et le maintien de la posture la mieux adaptée à la réalisation des mouvement volontaires.
Vue générale
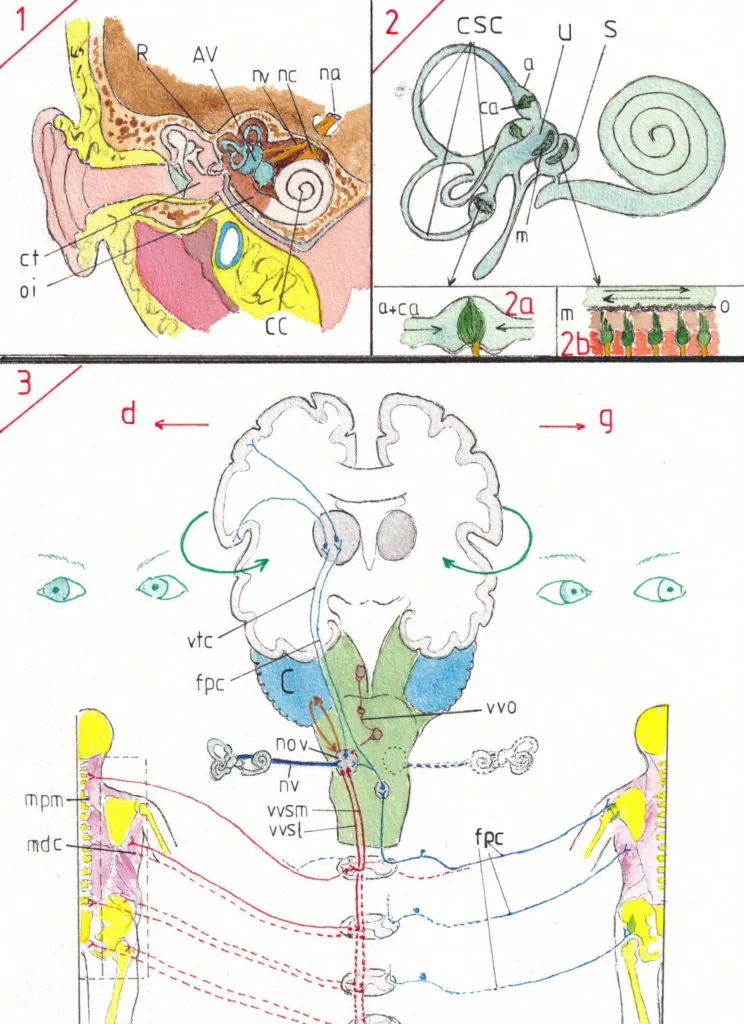
1- La voie vestibulaire
Elle est désignée classiquement sous le nom de voies (car elles sont au minimum quatre) vestibulo-spinales ( reliant l’appareil vestibulaire à la moelle épinière).
L’appareil vestibulaire (AV) -fig1- est situé dans l’oreille interne (OI) juste en dedans de la caisse du tympan (CT) en pleine épaisseur de l’os du rocher (R) ; il s’agit d’une formation complexe formant un tout avec le canal cochléaire (CC) lequel est dédié à la fonction auditive. De chacune de ces formations se détache un nerf sensoriel (nerf vestibulaire (nv) et nerf cochléaire (nc) pour former un nerf d’aspect unique nerf auditif (na) ou VIII ième paire des nerfs crâniens) ; ce dernier sort par un orifice osseux dans la cavité crânienne (fosse postérieure) et rejoint le tronc cérébral après un très court trajet.
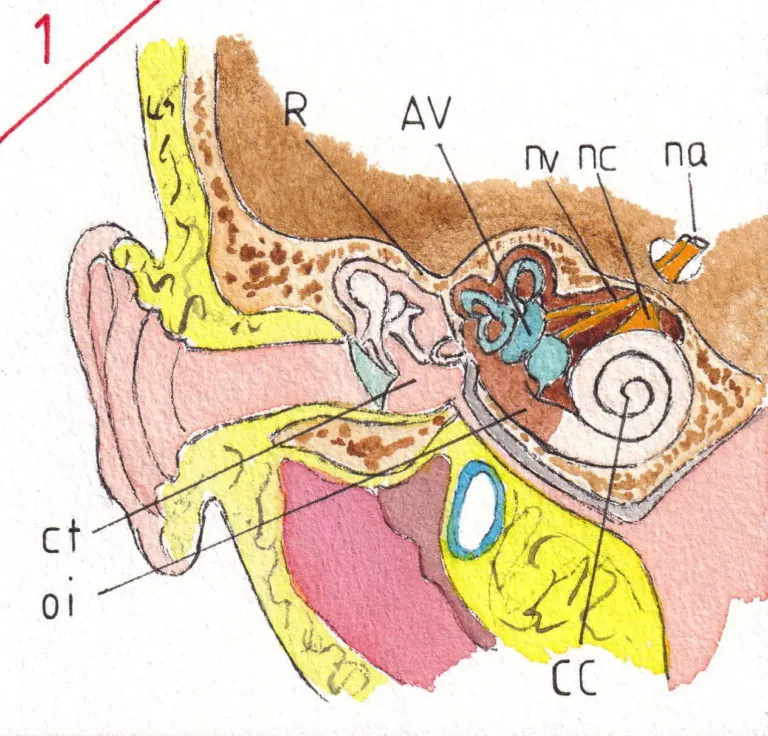
L’appareil vestibulaire (fig 2) est formé de 2 cavités grossièrement arrondies l’utricule (u) et le saccule (s) et de trois canaux semi-circulaires (csc) se terminant dans l’utricule , chacun par un renflement appelé ampoules (a).
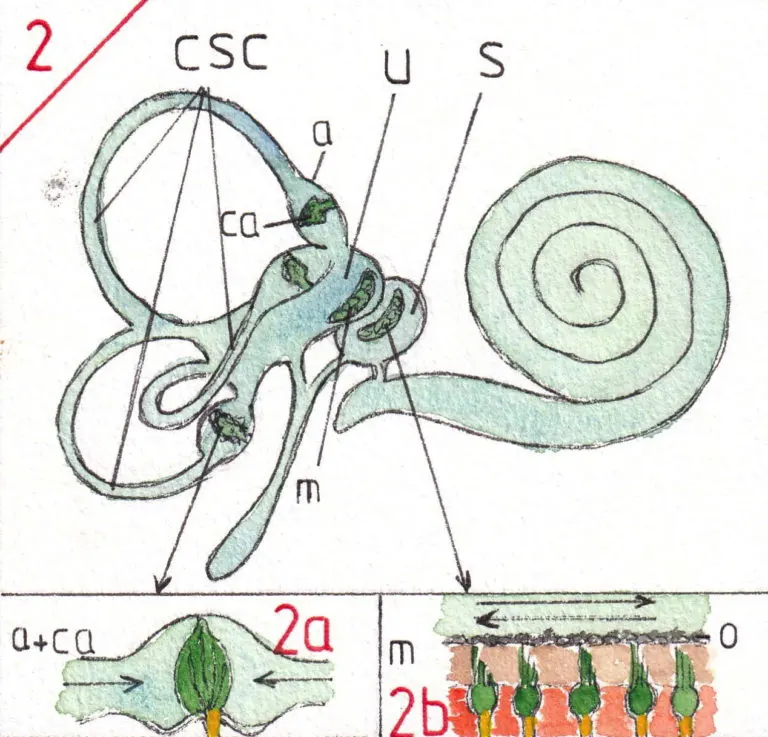
Cet ensemble est rempli d’un liquide dit endolymphatique et toutes les cavités communiquent entre elles ; le principe étant que le liquide se mette en mouvement à chaque mouvement de la tête ; les ampoules des canaux semi-circulaires (fig 2a) sont équipées de récepteurs en forme de crête dits crêtes ampullaires (ca) qui s’inclinent dans le sens du courant et sont donc sensibles aux mouvements de rotation de la tête – la position des canaux semi-circulaires leur permettant de fonctionner dans les trois plans de l’espace.
Les récepteurs utriculaires et sacculaires sont appelés macula (m) (fig 2b) ; elles sont sensibles à la position de la tête par rapport à l’horizontale car les cellules nerveuses qui y sont inclues tournent vers le liquide endolymphatique des cils de différentes longueurs englués dans une substance gélatineuse assez molle et recouverts par de toutes petites formations calciques ou otolithes (o) qui se déplacent sur la couche sous-jacente selon la gravité ; de sorte que l’utricule et le saccule sont dédiés à la sensation de la position de la tête dans l’espace ainsi qu’aux accélérations linéaires ; chacune de ces cellules nerveuses forment le point de départ d’une fibre nerveuse (en jaune) qui formera le nerf vestibulaire.
Cependant il ne s’agit là que d’une partie du système vestibulaire qui a pour fonction de renseigner le cortex sensoriel sur les mouvements et la position de la tête; son atteinte peut être très gênante mais finit souvent par être compensée par la deuxième partie du système vestibulaire centré sur le noyau vestibulaire (nov) ; en effet (fig3) , après avoir pénétré dans le tronc cérébral le nerf vestibulaire (nv) se termine dans un noyau situé à la jonction protubérance-bulbe et dont l’intégrité est d’ abord nécessaire aux transferts des informations sensorielles de l’appareil vestibulaire vers le cortex.
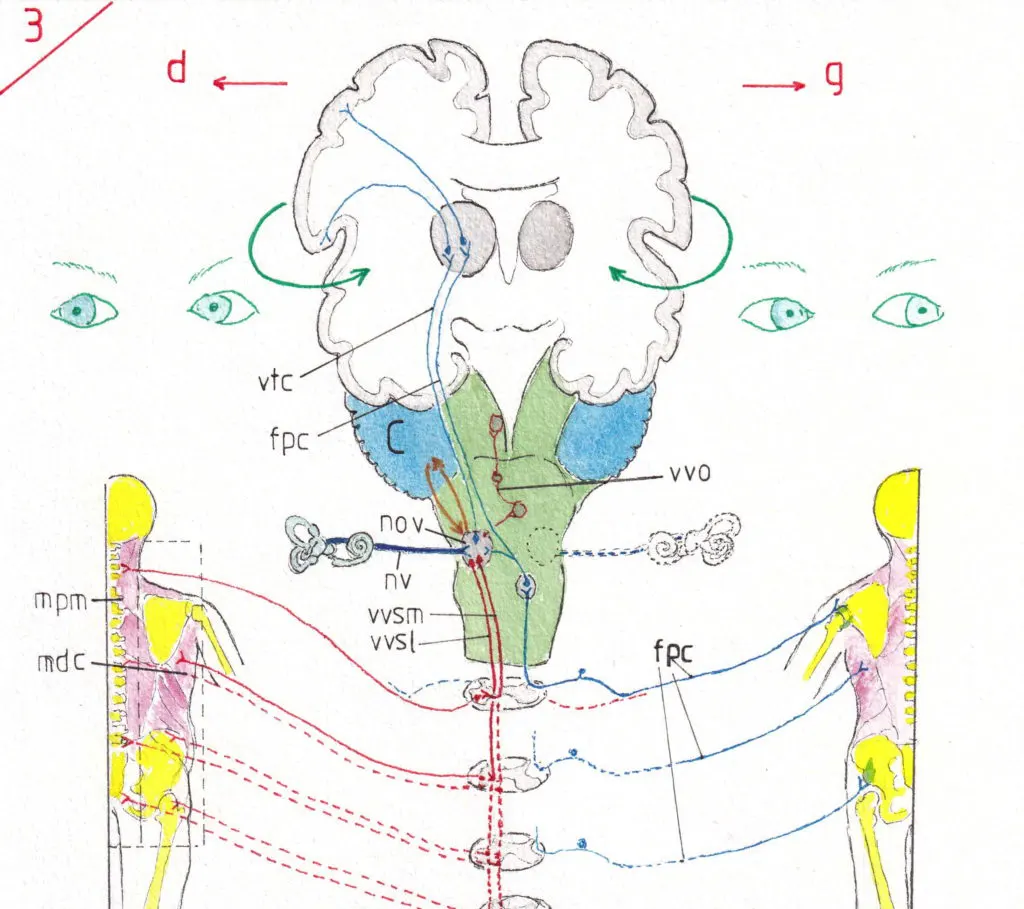
En effet des fibres de la sensibilité proprioceptive (en bleu sur le schéma) transmettent la sensibilité profonde des muscles, tendons et articulations provenant de l’hémicorps opposé (car passant la ligne médiane après relais dans leurs noyaux intermédiaires) ; ces fibres proprioceptives (fpc) ramènent donc toute la sensibilité profonde au cortex sensoriel d’un hémicorps avec un relais classique dans le thalamus mais auparavant elles font relais sur le noyau vestibulaire du côté opposé ; les noyaux vestibulaires sont donc renseignés avant le cortex de tout changement de position du corps et de chacun de ses segments ; il répond aussitôt par ses deux voies motrices (en rouge, 2 de chaque coté, en envoyant les corrections nécessaires aux différents muscles nécessaires à l’équilibration par les voies vestibulo-spinales médiales (vvsm) et latérales (vvsl) ; les premières sont destinées après relais dans les nerfs moteurs de la moelle aux muscles paramédians (mpm) dont le cou ; les secondes aux muscles des ceintures (mdc) épaule , tronc latéral , hanche , haut de cuisse ) ; ainsi ces voies vestibulo-spinales peuvent elles adapter le corps à tout changement de position ou d’équilibre avant que le cortex n’ait eu le temps de répondre ; de même en cas de chute ce sont elles elle qui mettront en œuvre dans l’extrême urgence la position de réception la moins risquée pour l’ensemble du corps . Notons que le schéma montre des voies sensorielles et motrices ne semblant fonctionner que d’un seul côté alors que les deux entrent en jeu bien entendu simultanément.
Le noyau vestibulaire commande aussi en grande partie le mouvement spontané des yeux en raison de ses relations automatisées entre lui et les noyaux moteurs oculaires voies vestibulo-oculaires (vvo) permettant au regard de continuer à viser une cible malgré un déplacement de la tête : si la tête tourne à gauche les yeux tournent à droite et inversement ; ce réflexe qui ne peut être modifié que par des influx volontaires corticaux sont à la base de la recherche clinique des nystagmus physiologique ou pathologique.
Pour terminer indiquons les importantes voies (courtes flèches en orange) qui relient le noyau vestibulaire au cervelet (C) qui seront vues avec la physiologie de cet organe.
2- La voie réticulaire
Concerne le transfert d’information de la formation réticulaire (FR) située dans le tronc cérébral vers la moelle épinière.
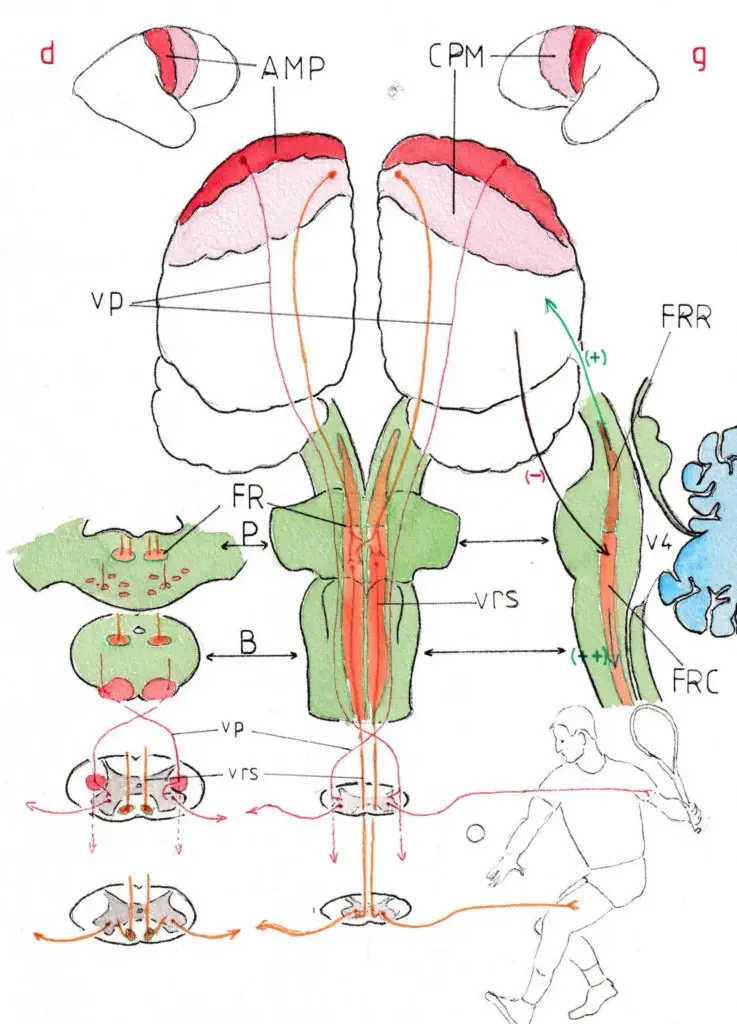
La formation réticulaire siège de chaque côté de la ligne médiane et s’étend depuis le haut des pédoncules cérébraux jusqu’à la partie basse du bulbe où la limite avec la substance grise de la moelle épinière devient imprécise ; elle présente par ailleurs une structure neuronale très complexe faite d’amas neuronaux plus ou moins bien individualisés et non de noyaux gris classiques faits de corps cellulaires regroupés; d’où son nom de réticulaire ( « semblable à un filet »). elle est classiquement séparée en deux parties aux limites également assez imprécises :
La partie haute (en marron sur les schémas ) correspond à la FR rostrale (FRR);
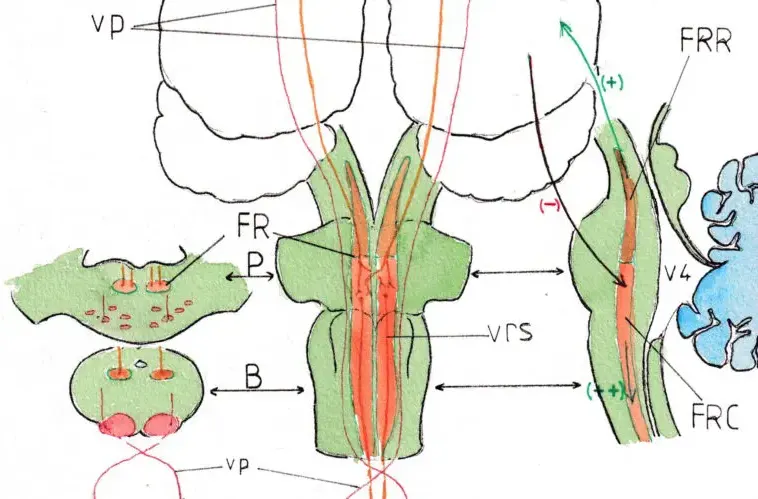
elle dirige ses connexions vers le cerveau sur lequel elle a une fonction classiquement activatrice (+) ce qui pourrait expliquer que son inactivation momentanée soit responsable du coma et sa reprise d’activité de l’éveil . Ses fonctions sont en réalité complexes; elle est en particulier considérée comme responsable des alternances veille -sommeil.
Formation réticulaire et voies réticulo-spinales
Seule la partie basse (FR caudale FRC) en orange nous intéresse dans cette page dédiée à la motricité ; celle-ci s’étend du milieu de la protubérance à la partie basse du bulbe (schéma central) ; les schémas en coupe horizontale de la protubérance (P) et du bulbe (B) (à gauche ) montrent sa situation bilatérale et postérieure ; celui de droite est une coupe paramédiane montrant la position de la SR tout le long du tronc cérébral devant les structures ventriculaire (V4) et canalaire. Cette partie caudale est nettement activatrice (++) mais tempérée par des influx inhibiteurs (–) venant de l’encéphale sus-jacent (tronc cérébral supérieur et cortex ). La libération de cette inhibition explique sans doute l’hypertonie et la spasticité survenant quelques semaines après une paralysie initialement flasque par lésion corticale ou tronculaire alors qu’elle est plus modérée dans les lésions médullaires.
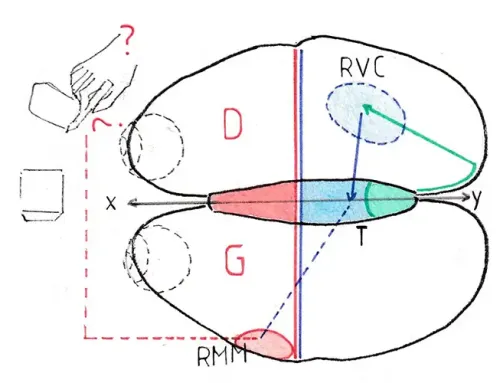
Nous avons tenté d’expliquer le fonctionnement physiologique de cette voie réticulaire par un exemple et la correspondance des schémas : le coté droit de l’encéphale se trouve à gauche (g) et inversement (d) puisque l’ensemble est vu de face avec des coupes étagées à droite:
La partie rouge du cortex représente l’aire motrice primaire (AMP); la partie rose le cortex prémoteur (CPM) dont nous avons vu le rôle majeur à la page » 1-Voies pyramidales » ; du cortex moteur (droit et gauche) partent les voies pyramidales (vp) sur lesquelles reposent les mouvements volontaires : en l’occurrence ici les mouvements des bras d’un joueur de tennis qui s’apprête à renvoyer une balle ; ces voies croisent la ligne médiane et le cortex moteur droit est particulièrement sollicité dans notre exemple puisqu’il va relancer la balle avec son bras gauche (le joueur est ici gaucher!) ; cependant pour que le coup soit réussi il ne suffit pas que le bras exécute parfaitement le geste , il faut aussi que l’ensemble du corps soit placé au bon endroit (probablement le système pyramidal volontaire a-t-il fait courir le joueur jusque -là !) mais les différents segments du corps doivent prendre une position idéale : les cuisses doivent fléchir, le tronc se tourner vers la gauche, les muscles qui actionnent cet ensemble pour certains se contracter , d’autres se relâcher ; certains doivent assurer la solidité de la position et prévoir que celle-ci peut se trouver compromise lors de la violente frappe à venir ; de toute cette activité musculaire extrêmement complexe le joueur n’a aucune conscience ; tout ce travail complémentaire est l’œuvre des voies réticulo-spinales (vrs en orange) dont l’activité a été mise en route avant celle des voies pyramidales dans le cortex prémoteur grâce à ses connexions avec les neurones de la SR ; ces voies assurent donc l’automaticité de la préparation et aussi celle de la bonne position et de la tonicité du corps lequel bénéficiera aussi de toute cette assistance jusqu’à la fin du déploiement du mouvement.
Dans la réalité les choses sont bien sur infiniment moins schématiques ; le cortex prémoteur n’est pas le seul à être connecté avec la SR ; le cortex moteur primaire l’est aussi et sans doute d’autres territoires corticaux ; par ailleurs les voies vestibulo-spinales décrites plus haut jouent aussi leur rôle lors d’un mouvement mettant en jeu l’équilibre du corps ; mais il faut insister sur toute la subtilité et la complexité que se partagent les différentes voies motrices , volontaires et involontaires avant d’évoquer celles involontaires également du cervelet (Motricité cervelet ). et souvent la difficulté pour le clinicien d’interpréter la cause et le pronostic de certaines séquelles motrices des cérébrolésions.