Encéphale et automatismes neuro-végétatifs
A – La formation réticulée
Composée d’amas neuronaux à la distribution assez aléatoire elle peut cependant être systématisée anatomiquement en une formation médiane égrainée en noyaux, une formation paramédiane bilatérale et symétrique et une formation latérale également bilatérale et symétrique.
La fig 1 représente à gauche la projection de la formation réticulée (FR) sur la face antérieure du tronc cérébral et à droite deux coupes, l’une au milieu de la protubérance (P), l’autre à la partie supérieure du bulbe rachidien (BR) ; ces coupes montrent que la FR est globalement postérieure dans le tronc cérébral ; incidemment la coupe passant par la protubérance montre la proximité du noyau du VI (noyau moteur oculaire externe) avec la réticulée paramédiane -frpm-)(voir son rôle dans les mouvements conjugués des yeux à la page sur l’oculomotricité) ; sur la représentation de face on voit que la réticulée poursuit sa route à travers les thalamus (TH) vers le haut et vers la moelle épinière (ME) vers le bas ; la réticulée médiane (frm) est formée de noyaux séparés dits du raphé ; la réticulée latérale (frl) est plus homogène mais contient aussi des agglomérats neuronaux formant des noyaux de contrôle végétatif (respiration et pression artérielle-cf infra-).

1 ° Rôle de la réticulée sur l’éveil
Ce rôle est déterminé par l’action de deux types de neuromédiateurs sécrétés par les neurones de la réticulée :
Ceux situés dans la réticulée latérale sont essentiellement des neurones à noradrénaline qui ont une influence excitatrice à la fois sur le cortex et sur la moelle ; on lui donne le nom de réticulée activatrice ; elle est à la fois responsable de l’état de veille mais aussi de la tonicité des muscles du corps ; la partie vraiment activatrice serait située essentiellement dans le mésencéphale -M- (ou pédoncules cérébraux) partie la plus haute du tronc cérébral ; la réticulée sous-jacente est plutôt inhibitrice ; mais le cortex activé par la réticulée activatrice inhibe en réaction cette même réticulée activatrice pour éviter son emballement : ainsi explique t-on l’hypertonie des membres en cas de lésions des voies motrices pyramidales quand elles se situent au dessus du mésencéphale (ce qui est le cas le plus fréquent des lésions induites par le traumatisme crânien ; en effet les lésions à ce niveau ne peuvent plus jouer leur rôle régulateur sur la réticulée activatrice) ; à l’inverse l’hypotonie des membres quand elles se situent au dessous (lésions basses du tronc cérébral et de la moelle), la réticulée activatrice étant désactivée.
Par ailleurs la substance grise de la moelle subit une excitation provoquée par les mouvements du corps, excitation qu’elle transmet à la réticulée activatrice et de là au cortex expliquant l’état d’éveil favorisé par l’activité physique.
La réticulée médiane est surtout sécrétrice de sérotonine, neuromédiateur impliqué dans le contrôle du stress et du sommeil ; les axones de ses neurones rejoignent les voies réticulées latérales et c’est donc l’ensemble de ces actions plus complémentaires qu’opposées qui se distribuent au cortex après être passées par deux voies (fig 2) :
- L’une principale traverse les thalamus (TH) dans une tranchée médiane dite intralaminaire -til- (fig 1), avant de gagner le cortex.
- L’autre secondaire passe contre l’hypothalamus (HTH) ; de là les arborescences des neurones noradrénergiques et sérotoninergiques gagnent la totalité du cortex (fig 3).



La ramification dans les thalamus avant l’expansion corticale semble avoir une importance dans le processus attentionnel ; en effet une stimulation d’une zone précise des thalamus est transmise à la partie correspondante du cortex et permet de centrer une attention spécifique sur tel ou tel sujet.
A l’inverse le coma qui succède immédiatement à une cérébrolésion, quand celle-ci est importante, est sans doute lié à un arrêt de la fonction de la formation réticulée ; il se caractérise, dans sa variété la plus profonde par une totale perte de conscience avec absence de réaction à tout stimulus qu’il soit vocal ou douloureux ; les paupières sont fermées ; le sujet semble dormir mais il n’est pas réveillable ; peut-être s’agit-il là d’une protection réflexe du tronc cérébral pour stopper au maximum l’activité neuronale au sein d’un cerveau contus comme un disjoncteur stopperait tout transfert électrique en cas de court-circuit ; même si il faut prendre cette comparaison avec beaucoup de prudence, elle a inspiré l’idée que chez un blessé cérébral sévère la mise sous coma artificiel, en utilisant des drogues adaptées pourrait compléter cette « mise au repos » du cerveau pendant la phase aiguë ; si la démonstration de son utilité n’a jamais pu être absolument prouvée, elle reste cependant utilisée de façon régulière lors de la prise en charge initiale ; si le coma artificiel (ou sédation profonde) peut être prolongé le temps voulu, le coma spontané, hormis les cas très graves, ne dépasse presque jamais trois semaines à un mois ; la sortie du coma se manifeste par une ouverture des yeux, mais pas forcément par une reprise de la conscience : pour reprendre la comparaison précédente le disjoncteur (la formation réticulée) est remis en marche mais si l’électricité est rétablie il faut aussi que les lampes ne soient pas trop abîmées ; ainsi la qualité de la reprise de la conscience dépend de la gravité des lésions cérébrales ; de plus il y a une relation certaine entre la durée du coma spontané et celle du retour de la conscience.
2° Rôle de la réticulée dans le contrôle de la tension artérielle.
Fig 4

La formation réticulée possède des noyaux de contrôle de la TA selon des modalités réflexes exposées succinctement ci -dessous ; cependant elle est loin d’avoir le rôle principal dans cette régulation, l’essentiel étant assuré par le système rénine-angiotensine d’origine rénale qui ne sera pas évoqué ici car elle ne concerne pas le cerveau.
Il existe à l’intérieur de la formation réticulée trois centres nucléaires groupés en un centre unique dit centre vaso-moteur (CVM) situé entre le tiers inférieur de la protubérance et les 2/3 supérieurs du bulbe rachidien ; deux sont des noyaux excitateurs situés de façon bilatérale et symétrique au dessus du troisième unique et inhibiteur ; les noyaux excitateurs (NE) ont une action tonique permanente sur la paroi musculaire lisse des vaisseaux sanguins et sont responsables du tonus vaso-moteur ; cette action s’effectue grâce au système sympathique ayant son système de commande étagé de l’hypothalamus à l’ensemble de la substance grise de la moelle épinière (pour plus de détails voir infra à hypothalamus).
Le centre inhibiteur (CI) est relié au système parasympathique dont le poste de commande se situe dans le noyau dorsal du nerf pneumogastrique-ndX- (un de chaque côté) ; son siège se trouve dans le bulbe rachidien. Le nerf pneumogastrique, ou nerf vague, dixième paire des nerfs crâniens (ou X) sort de la boite crânienne par le foramen jugulaire et se distribue à la majorité des viscères thoraciques abdominaux et pelviens sans passer par la moelle épinière ; mais il n’a pas d’action directe sur les vaisseaux périphériques car ses ramifications sont limitées à la sphère viscérale (voir infra) ; cependant son noyau de commande peut inhiber le centre de commande sympathique qui se trouve au contact de l’hypothalamus ; cette inhibition contraint le sympathique a cesser son action vasoconstrictrice ; le parasympathique devient donc vasodilatateur ; de plus les deux systèmes ont une action opposée sur le plan cardiaque : le sympathique accélère le coeur et augmente sa force de contraction ; le parasympathique fait l’inverse ; le premier est donc hypertenseur ; le second hypotenseur ; c’est sur cette base que s’effectue la régulation de la TA d’origine encéphalique.
D’abord le centre vasomoteur reçoit des influences corticales transmises par l’intermédiaire de l’hypothalamus ; l’effet peut être inhibiteur mais il est le plus souvent excitateur : l’ensemble du cortex est concerné avec une mention spéciale pour le cortex cingulaire, l’hippocampe et l’amygdale très impliqués dans les émotions : on connait la réaction classique de vasoconstriction sous-cutanée (pâleur) et d’accélération du rythme cardiaque (sensations de palpitations) qui accompagnent la colère ou la peur ; à l’inverse une émotion brutale peut stimuler anormalement le noyau dorsal du vague et provoquer un ralentissement cardiaque et une vasodilatation périphérique ,donc une hypotension avec quelquefois une perte de connaissance appelée malaise vagal.
Une exception à la vasoconstriction généralisée d’origine sympathique existe cependant lors de l’exercice musculaire : en effet les vaisseaux des muscles en action échappent à ce mécanisme de vasoconstriction pour leur permettre une augmentation de l’apport en oxygène par voie sanguine ; cet effet est lié à des boucles réflexes portées par le système sympathique médullaire.
Ces actions ponctuelles sur le centre vasomoteur sont en permanence contrôlées par le système réflexe des barorécepteurs ; ces derniers siègent à plusieurs endroits sur des gros vaisseaux sanguins mais plus particulièrement à l’origine des deux carotides internes (CInt). Ils sont formés de terminaisons nerveuses enchâssées dans le renflement musculaire de la paroi de ces artères à leur origine ; ces renflements sont appelés sinus carotidiens (SC) ; toute déformation de leur paroi provoquée par un changement brutal de la pression artérielle est captée par ces terminaisons et amenée par des nerfs spécifiques dépendants des nerfs glossopharyngiens (IX) qui conduisent l’information aux noyaux solitaires -NS- (communs aux VII , IX et X -voir page nerfs crâniens) et enfin au centre vasomoteur qui réagit en fonction des informations reçues ; toute baisse brutale de pression dans les sinus conduit à une vasoconstriction et à une augmentation du rythme et de la force de contraction cardiaque et inversement ; un exemple familier de ce type de réflexe peut être éprouvé lors d’un brusque passage de la position allongée ou assise à la position debout ; celle-ci chasse le sang vers le bas du corps et entraine une hypotension transitoire dans le territoire céphalique avec, chez le sujet âgé notamment, étourdissement passager et risque de chute ; cette baisse de tension doit être immédiatement compensée par de puissants influx venant des deux centres excitateurs bulbaires pour rétablir une bonne vascularisation cérébrale.
3° Rôle de la réticulée dans le contrôle de la respiration
(fig.5)

A la différence des automatismes cardiaques dont les centres nerveux de base se trouvent dans la paroi cardiaque elle-même, les automatismes respiratoires de base ont leurs centres dans le tronc cérébral ; ils se composent d’un centre médian dit pneumotaxique (cmpt) situé à la partie haute de la protubérance, de trois centres bulbaires : deux sont bilatéraux et symétriques : ce sont les centres expiratoires (cex) ; le troisième, le plus important, est le centre inspiratoire (cin) médian : c’est lui qui détermine le rythme de base et continue même de fonctionner en cas de dénervation complète. Le centre pneumotaxique ne fait que bloquer l’inspiration suivant le rythme imposé par les automatismes du moment ou par la volonté. Quant aux centres expiratoires ils n’interviennent vraiment qu’en cas d’expiration forcée.
Les modifications automatiques du rythme respiratoire sont adaptées aux besoins en oxygénation du sang, mais aussi et surtout à celui de l’élimination du gaz carbonique (CO2) suivant les circonstances physiologiques du moment, la plus courante étant l’adaptation à l’effort ; pour cela le système a de nombreuses ressources :
- Certaines sont prévisionnelles et d’origine corticale comme l’hyperventilation relative qui précède de peu un effort prévu alors qu’aucune modification métabolique n’a encore pu se produire et qu’elle n’a pas vraiment de raison d’être à cette étape. L’origine en est sans doute d’ordre émotionnel
- Une seconde, plus importante prend en compte les modifications des concentrations en oxygène du sang artériel circulant : elle agit par l’intermédiaire de chémorécepteurs artériels situés au niveau des bifurcations carotidiennes et sur la crosse de l’aorte, tout près des barorécepteurs vus au chapitre précédent et à ne pas confondre avec eux car leur mode d’excitation est très différent : ce sont de petites formations tissulaires vascularisées (ou glomus-g-) par une artériole en prise directe avec le courant artériel adjacent ; sur ces glomus sont implantés des récepteurs sensibles à la pression artérielle en oxygène ; les nerfs de transmission sont identiques à ceux qui véhiculent les informations de pression précédemment décrits ; ils rejoignent eux aussi les nerfs glossopharyngiens (IX) ; celles issues des corpuscules aortiques sont transportées par des branches des pneumogastriques (X) ; tous ces influx se terminent dans les noyaux solitaires et de là accèdent au centre respiratoire.
- Les propriorécepteurs (récepteurs tissulaires conscients et inconscients) situés sur les bronches, bronchioles, alvéoles pulmonaires sont reliés au noyau solitaire (NS) par le X assurant ainsi un contrôle sensitif d’origine pulmonaire.
- Le mécanisme de contrôle le plus important est situé sur l’aire chimio-sensible bulbaire (ACS) ; en effet le centre inspiratoire n’est vraiment stimulé que lorsque la baisse de la pression artérielle en oxygène tombe au dessous de 10% de sa valeur normale ce qui n’arrive que lorsque le ventilation chute énormément ; les chémorécepteurs artériels précédemment décrits ne sont donc que rarement stimulés ; le stimulant le plus important de la ventilation est en fait l’élévation du CO2 (gaz carbonique) : ceci est parfaitement logique puisqu’il est le produit final du métabolisme et qu’il doit être éliminé au plus vite par les poumons ; le CO2 diffuse facilement dans le système nerveux central et notamment dans le liquide céphalorachidien (LCR) provoquant par sa liaison avec l’eau (H2O) la formation d’ions H+ qui sont le stimulus électif de l’aire chémosensible (ACS) ; celle-ci se situe très près de la surface antérieure du bulbe et les ions H+ y diffusent très facilement ; l’information est immédiatement transmise au centre inspiratoire qui peut réagir avec une grande rapidité à la moindre élévation de la pression du CO2 dans le sang artériel.
Quant à la partie mécanique de la ventilation elle est assurée par les muscles intercostaux mais surtout par le diaphragme (D) ; en effet les poumons sont solidaires de l’ensemble de la cage thoracique grâce à la pression négative qui règne dans les plèvres et toute distension de ce contenant distant le contenu, en l’occurrence les poumons qui se remplissent d’air à l’inspiration et le rejette à l’expiration ; les mouvements du diaphragme peuvent être assimilés à ceux d’un piston qui s’abaisse (à l’inspiration) et remonte à l’expiration : seul le mouvement d’abaissement est actif ; il est assuré par les 2 nerfs phréniques (np) ; nerfs moteurs du diaphragme émanant des branches motrices des nerfs cervicaux C3, C4 et C5 ; ils s’étendent du cou aux coupoles diaphragmatiques en longeant les bords du coeur ; arrivés sur les coupoles ils pénètrent leurs parties musculaires striées comme les nerfs moteurs de n’importe quel muscle, la différence essentielle étant dans leur fonctionnement rythmique automatique délivré par le centre inspiratoire bulbaire, toujours associé à la possibilité d’un contrôle volontaire comme nous pouvons l’expérimenter très facilement.
B – L’hypothalamus
1° Situation
Teinté eu rouge sur les deux figures 6, l’hypothalamus (hy) se situe (fig.6 a) sur les parois latérales et la paroi inférieure du troisième ventricule (V3) ; limité en avant par la lame terminale (lt) qui joint la commissure antérieure (ca) au chiasma optique (cho), en bas par le plancher du diencéphale jouxtant la paroi inférieure du troisième ventricule, ainsi que son bas-fond au dessus de la tige pituitaire (-tp– qui conduit à l’hypophyse –h-) et cela jusqu’aux tubercules mamillaires (tm) qui en font partie ; ses limites postérieure et supérieure sont moins précises ; en tout cas comme son nom l’indique la limite supérieure ne dépasse pas celle du thalamus (th).
La fig. 6 b est une coupe frontale passant par la verticale (en pointillé) de 6a soit un peu en avant des tubercules mamillaires ; les pointillés horizontaux indiquent les correspondances de hauteur des différentes structures entre les deux schémas ; sur le schéma 6b on trouve au contact des parois du troisième ventricule un noyau qui l’entoure et correspondant au noyau peri-venticulaire (npiv) ; un peu en dehors ce sont les noyaux médiaux (nm) et plus en dehors encore les noyaux latéraux (nl) ; nous verrons plus loin que cette disposition est un peu différente en avant et en arrière de cette ligne ; au niveau ici choisi les noyaux médiaux peuvent être divisés en noyaux dorso-médiaux (ndm) et ventro-médiaux (nvm).

2° Relation de l’hypothalamus avec le système limbique
(fig7)
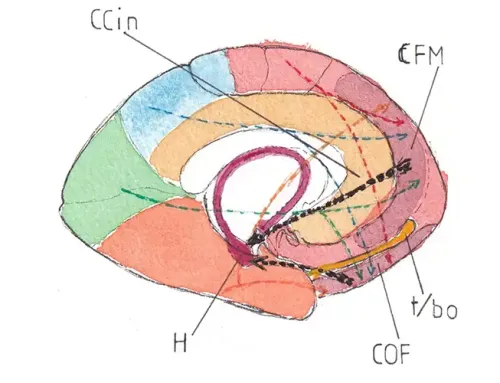
Se basant sur la neurophysiologie animale, le terme « limbus » ou « limite » a été donné à certaines structures de l’encéphale à une époque où l’on pensait qu’elles représentaient une limite fonctionnelle entre le système automatisé du tronc cérébral et le cortex hémisphérique (d’où le terme de système limbique) ; localisé au diencéphale il est apparu assez vite qu’on devait y intégrer les structures participant aux processus émotionnels et qui s’étendent relativement loin à la face interne des hémisphères ; ces structures ont toutes la particularité de converger vers l’hypothalamus qui en représente le centre, tant sur le plan fonctionnel qu’anatomique.

Les plus importantes sont :
- Au sein de la cinquième circonvolution temporale -T5- (ou gyrus parahippocampique ,l’amygdale (a) et l’hippocampe (hi) :
- l’amygdale (a) est responsable des sensations de peur ou du sentiment de punition pouvant évoluer selon les cas vers des réactions de fuite ou d’enragement ; mais étant composée de plusieurs noyaux la stimulation de certains peut aussi occasionner chez l’animal des comportements de plaisir. La destruction expérimentale des amygdales donne le syndrome de Klüver-Bucy très bien décrit chez le singe ; il peut se rencontre chez l’homme, quoique très rarement, la lésion bilatérale des amygdales étant exceptionnelle ; le syndrome comporte alors une oralité pathologique avec portée à la bouche de tout ce qui passe à proximité, une indolence totale avec absence de crainte et une hypersexualité.
- l’hippocampe (hi) est le centre de la mémoire épisodique (mémoire des évènements de vie). Les informations y sont traitées par un circuit complexe dit de Papez associant l’hippocampe, le trigone (tr), les tubercules mamillaires ™, la partie antérieure du thalamus (tha) et le cortex cingulaire (ccing).
- Le cortex olfactif (col) fait partie du système limbique ; de grande importance chez nombre d’animaux où il joue un rôle majeur dans la discrimination alimentaire et dans la sexualité ; sa liaison étroite avec les noyaux hypothalamiques redouble son efficience.
- Le cortex cingulaire (ccing) (autour du corps calleux) fait partie en intégralité du système limbique ; nombre de ses fonctions restent cependant mal connues.
- Le cortex cingulaire antérieur (ccinga) a été le plus exploré ; on sait qu’il contrôle les émotions, l’attention et la mémoire ; mais aussi la motivation dans le but d’obtenir un résultat souhaité ; la déception quand le but n’est pas atteint ; au point d’être très suspecté d’avoir une responsabilité dans les idées négatives de la dépression ; il semble par ailleurs pouvoir comparer plusieurs choix possibles dans une prise de décision ; soit à l’avance, soit pendant l’action et permettre ainsi un changement d’orientation pour obtenir une solution plus satisfaisante ; en ce sens ses liaisons dans les deux sens avec la sensibilité émotionnelle de certains noyaux de l’hypothalamus paraissent très importantes.
- Le cortex cingulaire postérieur (ccingp) est moins connu ; il parait cependant très impliqué dans la récupération de la mémoire épisodique (il se trouve atteint préférentiellement dans la maladie d’Alzheimer) ; mais son rôle ne s’arrêterait pas là : il aiderait lui aussi aux prises de décision en évaluant les expériences passées et en utilisant les composants émotionnels qui y sont associés ; son action est forcément importante car c’est une des régions corticales les plus actives métaboliquement lorsque l’on utilise des explorations fonctionnelles (IRM fonctionnelle et PET -SCAN).
- Enfin la projection de la formation réticulée (fr) dans l’hypothalamus peut être incluse dans le système limbique eu égard à l’importance des liaisons entre ces deux formations ; liaisons jouant dans les deux sens : tronc cérébral -cortex et cortex-tronc cérébral.
3° Hypothalamus et systèmes sympathique et parasympathique
(fig 8 et fig 9)
Les systèmes sympathique et parasympathique contrôlent par voie réflexe une grande partie du système autonome encore appelé système végétatif, car seul en activité en cas d’atteinte totale de la conscience comme on peut le voir chez les cérébrolésés en état végétatif ; certains organes ont une autonomie propre (coeur, reins, intestins, foie, etc…) ; d’autres ne sont autonomes que s’ils sont stimulés par des centres du tronc cérébral (c’est le cas de la respiration et de ses liaisons avec les centres respiratoires, comme vu plus haut ; autonomes ou non la plupart des organes nécessitent des adaptations permanentes provoquées par les changements de condition du corps et/ou du psychisme : les systèmes sympathique et parasympathique répondent à ces derniers critères ; par ailleurs ils jouent un rôle non négligeable dans la vie consciente car le psychisme, notamment émotionnel, a une grosse influence sur leur activité et les réactions périphériques qu’ils provoquent sont souvent ,dans ces conditions, ressenties consciemment.
Ils ont le plus souvent des fonctions opposées, mais pas toujours, et leur action finale est en général la recherche du meilleur équilibre pour le bon fonctionnement des organes qu’ils innervent et cela en général à l’insu de notre conscience ; cette dernière n’étant alertée qu’en cas de dépassement de l’activité de l’un ou de l’autre : malgré leur rôle de régulateur il existe donc des pathologies de leur fonctionnement.
Enfin les relais des informations qu’ils véhiculent suivent des voies très différentes : hormis quelques noyaux parasympathiques crâniens à destinée céphalique le parasympathique circule dans le corps totalement en dehors de la moelle épinière par la voie des deux nerfs vagues (ou pneumogastriques ou X) alors que la voie sympathique utilise exclusivement la moelle épinière ; ses filets nerveux en sortent de haut en bas avec les nerfs rachidiens auxquels ils sont assez intimement mêlés.
La fig 8 montre les rapports des deux voies avec l’hypothalamus (hy) dont on connait déjà le rôle essentiel dans le relais des émotions qui se créent en permanence dans le système limbique ; on verra plus loin que les noyaux de l’hypothalamus ont eux-mêmes une grande importance dans la genèse de certaines émotions ; il existe de façon indiscutable un centre sympathique (cΣ) à l’intérieur de l’hypothalamus précisément au niveau du noyau postérieur qui est une aire assez vaste en contact étroit avec la formation réticulée qui remonte jusqu’à lui ; le centre parasympathique (cpΣ) n’est pas localisé de façon aussi précise ; paraissant situé aussi en région postérieure il n’est pas signalé par tous les auteurs mais on voit mal comment certaines émotions pourraient activer les nerfs vagues de façon souvent spectaculaire sans un relai à ce niveau ; la fig 8 n’est donc qu’un schéma très approximatif et surtout simplifié des voies nerveuses Σ (vΣ) et paraΣ (vpΣ) circulant entre l’hypothalamus et leurs noyaux particuliers :

- en vert la voie paraΣ se distribue aux noyaux des III , VII , IX et X ; les noyaux para Σ des X sont appelés noyau dorsal du vague (ndv) et sont le point de départ de l’innervation paraΣ du reste de corps par l’intermédiaire des pneumogastriques; comme déjà vu ils quittent la base du crâne par les foramens jugulaires de chaque côté du trou occipital .
- En orange les voies sympathiques n’ont pas de noyaux dans le tronc cérébral mais uniquement dans la moelle épinière à chaque métamère (cad à chaque espace intervertébral), leur sortie de la moelle se faisant avec le nerf rachidien correspondant ; voir infra.
La fig 9 montre le cheminement des deux voies (Σ en orange ; paraΣ en vert) à destination des organes cibles ; à la droite du schéma se trouve une demi-vertèbre et une demi-moelle en coupe horizontale ; à gauche et en haut les cibles musculaires et sous cutanées (msc) qui n’intéressent que le Σ ; en bas la partie viscérale (vis) (ici on a pris l’exemple d’un fragment d’intestin) qui intéresse les deux voies ; sont figurés en pointillés les changements de hauteur des nerfs par rapport à leur sortie du tronc nerveux (bulbe pour le X et segment de moelle pour le Σ) et l’organe cible ; ainsi le X est représenté en pointillé car descendant verticalement du bulbe ; il n’est figuré en trait plein qu’à la hauteur de l’organe cible ; ceci est important car les ganglions dont nous allons parler se trouvent presque toujours au dessous de la sortie correspondante des nerfs de l’axe nerveux.

Les nerfs sympathiques (en orange) naissent dans la corne latérale de la moelle (clm), alors que les nerfs moteurs somatiques (en rouge) naissent de la corne antérieure et que les nerfs sensitifs somatiques (en bleu) rejoignent la corne postérieure avec leur noyau extra-médullaire formant le ganglion spinal (gs) (voir pages consacrées à la motricité et à la sensibilité du corps).
Sont figurés ici deux neurones Σ, l’un destiné aux viscères (vis), l’autre aux muscles et à la région sous-cutanée (msc) ; les deux neurones se dirigent vers une formation ganglionnaire située devant la face antérieure et latérale de la vertèbre considérée et appelé ganglion sympathique (g Σ) ; dans le ganglion le neurone destiné aux viscères ne fait que passer et descendra plus bas vers une formation plus volumineuse appelé plexus (Px), également situé en prévertébral, dans lequel il fera relais avec un neurone de deuxième ordre qui se terminera sur l’organe concerné ; le neurone destiné aux muscles et à la région sous cutanée fait relais dès le ganglion sympathique et le neurone de deuxième ordre rejoindra son nerf rachidien d’origine pour innerver les régions musculaires et sous-cutanées concernées par ce même nerf rachidien.
Les neurones paraΣ (en vert) transitent par les nerfs pneumogastriques (X), totalement en dehors de la moelle épinière mais passent par les plexus dans lesquels ils ne font pas relais ; les neurones de deuxième ordre très courts naissent très près de l’organe concerné ou à l’intérieur même de sa paroi.
Les muscles et la région sous-cutanée (msc) ne sont donc innervés que par le Σ ; ses actions principales sont la vasoconstriction sous-cutanée (vcsc) et la sécrétion des glandes sudoripares (sgs) expliquant la pâleur et la sudation qui accompagnent souvent le stress ; sur le plan musculaire il provoque plutôt une vasodilatation artérielle (vdam) notamment à l’effort pour augmenter l’oxygénation des muscles en action (déjà signalé plus haut).
Les viscères (vis) reçoivent une double innervation Σ et paraΣ qui s’articule avec l’innervation et/ou la musculature intrinsèques des organes considérés ; l’action des deux systèmes est le plus souvent antagoniste car les neurones de deuxième ordre sécrètent des neuromédiateurs différents (adrénaline en général pour le Σ et acétylcholine pour le paraΣ) et d’action opposée ; mais l’action de ces neuromédiateurs peut varier aussi suivant la présence sur l’organe cible de neurorécepteurs variés (voir page sur la transmission de l’influx nerveux) induisant des effets d’apparence paradoxale.
Nous signalons pour terminer quelques effets très classiques de ces deux systèmes sur trois organes particulièrement vitaux :
coeur : le Σ accélère le coeur et augmente sa force contractile ; pour les artères coronaires il peut être vasodilatateur ou vasoconstricteur suivant les récepteurs ; le paraΣ ralentit le coeur, diminue sa force contractile et dilate les coronaires.
poumons : le Σ dilate les bronches ; le paraΣ les contracte ; nous voyons là l’utilité des médicaments dits sympathomimétiques (qui ont des effets voisins de l’adrénaline) lors de crises graves d’asthme.
tube digestif : le Σ ralentit le péristaltisme intestinal et renforce le tonus des sphincters ; le paraΣ fait l’inverse ; le premier ralentit donc le transit alors que le second l’accélère.
Nous arrêtons là cette courte liste qui nous éloigne un peu trop de l’encéphale ; elle est en effet assez longue et peut-être trouvée facilement en tableaux récapitulatifs sur tous les précis de physiologie ; nous avons fait ce détour pour montrer à quel point l’ensemble de l’organisme est associé à ce qui se passe dans l’encéphale ; les deux chapitres suivants vont nous en convaincre encore davantage.
Pour terminer signalons que les deux systèmes sont pourvues d’une sensibilité viscérale ramenant les influx vers l’encéphale ; ils sont représentés en doublons noirs sur le schéma ; ceux faisant partie du Σ (ssΣ) remontent avec les nerfs correspondant jusqu’à la moelle épinière puis l’encéphale ; ceux du paraΣ (sspΣ) avec les nerfs X jusqu’aux deux noyaux sensitifs bulbaire des X : les noyaux solitaires (v. nerfs crâniens). Ce sont toutes ces afférences sensitives qui sont à l’oeuvre dans les douleurs d’origine viscérale.
4° Les noyaux de l’hypothalamus
( fig 10)

L’hypothalamus a bien d’autres rôles que d’être un simple intermédiaire entre le cerveau et le corps ; il joue en effet des rôles spécifiques dans beaucoup de domaines, soit seul, soit associé à des structures déjà décrites notamment dans le domaine comportemental tant les effets y paraissent souvent complémentaires avec celles des formations adjacentes.
Ces actions sont liées à un assez grand nombre de noyaux, du moins vu la petite taille anatomique qu’il occupe (voir fig1) ; bien que l’isolement précis de ces noyaux, très proches les uns des autres, soit difficile et qu’il persiste encore certaines imprécisions dans leur représentation nous tentons d’en donner une description à la fois simplifiée mais assez communément admise, ainsi que les fonctions qui les accompagnent tout en gardant en tête que beaucoup de recherches sur l’hypothalamus ont été faites sur l’animal et ne sont pas forcément entièrement transposables à l’homme ; cependant de nombreuses situations pathologiques et leurs causes anatomiques ont prouvé le bien-fondé de la démarche.
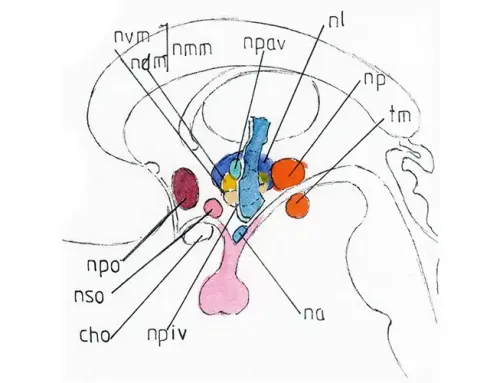
On peut classer ces noyaux en antérieurs, moyens et postérieurs dans le sens antéropostérieur, et périmédiaux, paramédiaux, médiaux et latéraux dans le sens transversal.
Noyaux antérieurs
Le noyau préoptique (npo) est le plus antérieur (quelquefois représenté en deux noyaux séparés -médial et latéral-) ; il se trouve juste en arrière de la lame terminale (lt) ou à cheval sur elle ; son excitation provoquerait plutôt une inhibition globale et une somnolence ; mais il est surtout notre « thermostat portatif » par lequel se règle en permanence notre température corporelle autour de 37°. Une explication de la fièvre en cas d’infection serait sa grande sensibilité aux substances pyrogènes issues notamment des toxines microbiennes ; il réagirait via le système sympathique par une vasoconstriction périphérique limitant la perte calorique et une augmentation du métabolisme de base ; et via la substance réticulée par une hypertonie musculaire avec contraction rythmique lors du frisson permettant, par l’augmentation de l’activité musculaire un réchauffement rapide ; la sensation de froid disparaitrait lorsque les deux températures (thermostat et corps) s’équilibrent ; la disparition de la cause pyrogène captée par le thermostat le ramènerait à la baisse ; son action serait alors d’inhiber totalement le sympathique provoquant une vasodilatation sous-cutanée (rougeur diffuse) relayée par une sudation intense (celle-ci non de nature sympathique) et permettant au corps le rétablissement d’une température normale.
Plus bas et un peu en arrière au dessus du chiasma optique (cho) se trouve le noyau supra-optique (nso) dont le rôle est surtout la neurosécrétion de la vasopressine -vsp (ou hormone antidiurétique ; v infra)
Noyaux moyens
- Le noyau périventriculaire (npiv) double les limites du troisième ventricule (v.fig1) ; il est représenté partiellement sur le schéma comme un segment de cette paroi ; très fin et large il est à la fois le point de départ de certains « facteurs de libération » (RH) à destination de l’antéhypophyse (v.infra) et semble jouer un rôle dans la régulation comportementale, son excitation provocant sentiment de déplaisir voire de rage.
- Le noyau paraventriculaire (npav) situé juste en dehors mais assez haut et petit est responsable de la sécrétion d‘ocytocine (ocy) (v.infra).
- Le noyau moyen et médial (nmm), un peu plus externe représenterait en fait deux noyaux : un supérieur appelé dorso-médian (ndm) : son excitation provoquerait une sensation de peur (comme pour l’amygdale) et un sentiment de punition, mais aussi certaines pulsions sexuelles ; un inférieur ou ventro-médian (nvm) serait celui du sentiment de récompense et de la satiété.
- Le noyau latéral (nl) (aussi appelé aire latérale) ; il est le noyau de la faim et de la soif et peut être en cause dans certaines formes (les plus rares) d’obésité ; il serait aussi celui dont l’excitation augmenterait l’activité globale physique et psychique ; par ailleurs suivant les degrés de stimulation il peut donner un sentiment de récompense ou au contraire d’énervement et de rage.
- Le noyau arqué (na) situé plus bas, découvert plus récemment semble être le point de départ de la sécrétion de certains « facteurs de libération » RH destinés à l’antéhypophyse (voir infra).
Noyaux postérieurs
- Essentiellement représenté par le noyau postérieur -np – (ou aire postérieure) il est sûrement le centre encéphalique du système sympathique ; sa stimulation augmente l’activité sympathique globale ; sa mise au repos la stoppe ; il entre en étroite connexion avec la formation réticulée dont le pôle supérieur l’entoure ; et c’est sans doute par lui que le centre thermostatique agit, du moins dans la régulation de la fièvre comme vu précédemment.
- Le tubercule mamillaire ™ très bien individualisé anatomiquement semble avoir les mêmes fonctions et lui appartenir.
5° Hypothalamus et hypophyse
(fig 11)

L’hypophyse est une glande endocrine (cad dont les sécrétions se font directement dans le sang pour être diffusées à l’ensemble du corps) la plus importante de notre corps ; elle se situe à l’intérieur de la boite crânienne, dans la selle turcique, dépression centrale de l’os sphénoïde ; elle est donc centrale et médiane ; cependant elle n’appartient pas totalement à l’encéphale car elle est formée de deux parties : une postérieure (ou post-hypophyse) qui est bien d’origine encéphalique ; l’autre antérieure (ou anté-hypophyse) est d’origine pharyngée ; ces deux origines se réunissent pendant la vie embryonnaire pour former une structure anatomique unique mais n’ayant pas les mêmes fonctions ; cependant l’intégration de l’antéhypophyse au système nerveux et en particulier à l’hypothalamus est complète ce dernier dirigeant en grande partie sa sécrétion ; l’ensemble ne peut donc être séparé ni anatomiquement ni physiologiquement.
D’abord ces deux glandes (antérieure et postérieure) sont toutes les deux sécrétrices d’hormones ; ensuite toutes les deux ont leurs noyaux de commande (soit directement pour la post.h- soit indirectement pour l’anté.h-) dans les noyaux hypothalamiques.
La post-hypophyse (PH) sécrète des neurohormones cad des hormones fabriquées par des neurones (ce qui est une exception) dont les corps cellulaires se trouvent dans l’hypothalamus (notamment dans le noyau supraoptique pour la vasopressine et dans le noyau paraventriculaire pour l’ocytocine) ; elles sont conduites par leurs axones dans la post-hypophyse ; de là un système capillaire les prend en charge pour les conduire dans la circulation générale :
- la vasopressine (vsp) (ou hormone antidiurétique -ADH-) cible le rein dans lequel elle provoque en continu la réabsorption de l’eau de l’organisme qui serait évacuée en quantité beaucoup trop importante en son absence ; son insuffisance de sécrétion est à l’origine du diabète dit insipide marquée par une diurèse très excessive et source de graves complications ; cette pathologie se rencontre parfois dans les suites précoces du traumatisme crânien ; elle y est le plus souvent temporaire.
- l’ocytocine (ocy) : son rôle principal est de provoquer l’éjection du lait des alvéoles de la glande mammaire pendant l’allaitement ; son rôle secondaire est de favoriser la contraction du muscle utérin pendant l’accouchement ; si cette deuxième action est physiologiquement assez peu importante, l’ocytocine en injection médicamenteuse est très utilisée en obstétrique pour déclencher le travail.
L’antéhypophyse sécrète 6 sortes d’hormones ; il s’agit soit d’hormones à action directe sur un organe cible, soit d’hormones provoquant la sécrétion hormonale d’autres glandes endocrines ; l’hypothalamus reste cependant un régulateur majeur de la sécrétion antéhypophysaire ; en effet ses neurones dont les corps cellulaires se trouvent surtout dans les noyaux paraventriculaire, periventriculaire et arqué secrètent encore des neurohormones, mais à action indirecte, dites « facteurs de libération » ou « releasing hormones » ou « RH » ; ces neurohormones sont libérées dans un système vasculaire (sv) situé dans et devant la tige pituitaire (tige de raccordement de l’hypophyse à l’hypothalamus) ; il s’agit d’un riche plexus vasculaire qui capte les RH et les transporte par voie veineuse (appelée système porte –sp-) dans l’antéhypophyse ou ils vont stimuler de façon élective les cellules glandulaires qui leur correspondent ; ces cellules glandulaires fabriquent leurs hormones spécifiques qui rejoindront la circulation générale à destination de leurs cibles : il ne s’agit plus là de sécrétion neurohormonale mais bien du mécanisme de sécrétion habituelle par des cellules glandulaires, comme cela se produit dans toute glande endocrine ; cependant l’originalité reste pour certaines hormones d’entre elles de contrôler la sécrétion d’autres glandes endocrines qu’elles ont pour cible ; en effet l’élévation comme la baisse des hormones des glandes endocrines cibles reviennent informer par voie sanguine le système hypothalamo-hypophysaire qui, comme un véritable chef d’orchestre peut augmenter ou diminuer les sécrétions des glandes cibles pour maintenir dans le sang le taux optimal de telle ou telle hormone à tel ou tel moment. C’est le mécanisme du bio feed-back.
Les 6 hormones antéhypophysaires sont :
- La TSH -tsh- (contrôlée par le TRH hypothalamique) est l’hormone antéhypophysaire qui stimule la glande thyroïde laquelle sécrète la thyroxine ; hormone dont le rôle principal réside dans la stimulation du métabolisme général.
- L’ACTH -acth- (contrôlée par le CRH hypothalamique) stimule la sécrétion du cortisol par les glandes surrénales ; le cortisol par de nombreuses actions périphériques favorise la résistance globale de l’organisme.
- Les hormones gonadotropes -ght- correspondent à deux hormones LH et FSH contrôlées par le GnRH hypothalamique ; chez l’homme la FSH stimule la production des spermatozoïdes, la LH active la sécrétion de testostérone par les testicules ; chez la femme la FSH stimule la production d’oestrogènes et la LH de progestérone par les ovaires.
- La GH –gh- ou hormone de croissance contrôlée par le GHRH hypothalamique est une hormone anabolisante majeure intervenant à la fois sur la croissance staturale et sur celle des organes profonds.
- La prolactine -pr-contrôlée par un PRH hypothalamique encore non formellement identifié stimule la croissance de la glande mammaire et la production de lait pendant la grossesse ; à l’inverse des autres hormones antéhypophysaires sa sécrétion est inhibée par l’hypothalamus en dehors de la grossesse ; la succion du mamelon entretient par contre sa sécrétion pendant la période d’allaitement maternel.
Ces indications sont données pour aider à mesurer les conséquences que des pathologies encéphaliques peuvent engendrer sur le système hormonal de l’ensemble du corps ; pour les lecteurs intéressés par ces sujets seule une étude attentive du fonctionnement de ces différentes glandes endocrines peut fournir des renseignements utiles et leur permettre de comprendre certaines pathologies induites par certaines affections neurologiques ou psychiques.
En dehors de ces actions hormonales maintenant connues depuis longtemps mais pour rester dans le domaine de l’encéphale il faut signaler la découverte assez récente d’une hormone sécrétée par le tissu adipeux et appelée leptine ; cette hormone agit sur le noyau arqué de l’hypothalamus lequel réduit la prise alimentaire et déclenche la puberté en stimulant le GHRH ; beaucoup de travaux ont été publiés sur l’insuffisance de sa sécrétion ou de l’inefficacité de ses récepteurs dans les mécanismes pathologiques aboutissant à certaines formes d’obésité.