Audition
A – Les voie auditives du pavillon au nerf auditif (ou mieux nerf cochléaire)
fig 1, 2, 3 et 4

1 – Le pavillon de l’oreille (P) conduit les sons vers la conque (C) sorte de renfoncement du pavillon qui tire son nom de sa ressemblance avec l’ouverture d’une coquille ; la forme de ces deux structures a, semble-t-il, une importance dans la sélection de certaines fréquences tonales
2 – Le conduit auditif externe (CAE) sert de chambre d’amplification de certains sons notamment ceux dont la fréquence est proche de celle de la voie humaine dont la perception est ainsi privilégiée ; par ailleurs ce phénomène explique en partie pourquoi les sons de très forte intensité de ce niveau de fréquence sont aussi les plus dangereux pour les structures très fines de l’oreille interne.
3 – Le tympan (ty) est une membrane fine et résistante, maintenue dans un état de tension adéquate par son adhérence au manche du marteau (mm) lui permettant de vibrer à la fréquence de propagation des ondes sonores incidentes ; elle est inclinée vers le bas et le dehors ; sur le schéma elle est présentée coupée verticalement pour découvrir la totalité de l’oreille moyenne ; elle marque la limite entre l’oreille externe et l’oreille moyenne ou caisse du tympan (CDT).
4 – La caisse du tympan (fig 1 et 2)

La caisse du tympan est une cavité creusée dans l’os du rocher et qui contient les trois osselets (Marteau M, enclume En et étrier Et) chargés de transmettre les vibrations sonores à l’oreille interne, point de départ de la stimulation sensorielle.
La caisse du tympan présente aussi à sa base l’ouverture d’un conduit très important : la trompe d’Eustache (TE) qui la met en relation aérienne avec la cavité du pharynx (PH) -cf infra-
Les osselets sont fixés entre eux par des ligaments assurant la continuité de la chaine mécanique qui se termine par la platine de l’étrier (Pet) laquelle s’adapte parfaitement avec la fenêtre ovale (FO), orifice de la face interne de la caisse du tympan et marquant la limite entre oreille moyenne et oreille interne ; la platine transmet les vibrations reçues sur le tympan au liquide peri-lymphatique (lpl) contenu dans l’oreille interne ; en effet elle s’enfonce dans le liquide à chaque onde transmise comme un micro-piston et répond ainsi précisément à la fréquence et l’amplitude des vibrations reçues par la chaine ossiculaire.
En plus des ligaments qui les fixent les uns aux autres, le marteau et l’étrier sont mis en tension par deux petits muscles : le tenseur du tympan (tdt) pour le premier et le muscle de l’étrier (mde) pour le deuxième (en pointillés sur la fig 2) ; non seulement ils assurent une tension constante sur le tympan afin que celui-ci soit toujours prêt à répondre de façon optimale, mais de plus ils protègent le système auditif de certains risques : des sons trop forts entrainent une hyper-contraction du muscle tenseur du tympan qui fixé au manche du marteau l’attire en arrière et bloque sa mobilité ; le muscle de l’étrier tend ce dernier vers l’arrière et le dehors limitant la course de la platine : en effet s’il se relâche (ce qui arrive souvent en cas de paralysie du nerf facial qui l’innerve) des sons même faibles peuvent être rendus plus forts et douloureux par un bombement excessif de la platine vers l’oreille interne ; ces deux muscles sont seulement suggérés en pointillés verts pour ne pas surcharger le dessin.
La trompe d’Eustache est une structure également très importante car sa perméabilité permet d’équilibrer en permanence les pressions atmosphériques qui règnent de chaque côté du tympan ; on peut en faire l’expérience très simple quand on monte en altitude où la pression atmosphérique baisse du côté du conduit auditif externe alors qu’elle reste plus élevée dans la caisse du tympan, ce qui limite la mobilité de celui-ci ; une simple manœuvre de déglutition qui ouvre le passage de l’air ambiant vers la caisse du tympan rétablit ainsi une pression identique de chaque côté du tympan qui retrouve ainsi sa mobilité optimale.
5 – L’oreille interne
Les fig 2 et 3 montrent la jonction de l’oreille moyenne avec la partie initiale des labyrinthes osseux et membraneux dédiés à l’équilibration (déjà étudié avec la motricité) et à l’audition formant l’oreille interne ; circulent dans ces labyrinthes le liquide périlymphatique (lpl) périphérique et le liquide endolymphatique (lel) central occupant le labyrinthe membraneux ; bien qu’il y ait des communications liquidiennes entre ces deux structures sensorielles (audition et équilibration), comme le montre le schéma, l’oreille interne fonctionne en totale indépendance : toute pression transmise à la platine se communique (flèche rouge du haut) au lpl de l’oreille interne situé en vis à vis, qui se trouvant repoussé, forme une onde liquide dans le premier espace ou rampe vestibulaire (RV) ; en théorie après passage sur toute sa longueur (3,5 cm de long pour les trois tours de spire) l’onde revient par un espace symétrique, la rampe tympanique –RT– (flèche rouge du bas) vers la fenêtre ronde (FR) occluse par une membrane assez lâche dont le bombement vers l’intérieur de la caisse du tympan stoppe le mouvement ; en effet les deux rampes communiquent par leur sommet (voir infra). Entre les deux rampes se trouve la rampe cochléaire (RC) remplie de liquide endolymphatique et sans communication avec la périlymphe.

La fig 3 montre la totalité de l’oreille interne ou cochlée : elle forme une sorte de colimaçon ; sa partie la plus à gauche sur le schéma correspondant à la fig 2 qui représente une vue en coupe passant à travers les deux fenêtres, ronde (FR) et ovale (FO) ; une large ouverture a été pratiquée sur la face antérieure de la cochlée pour en découvrir la structure interne et notamment ses trois rampes (les couleurs correspondent à celles de la fig 2) : la rampe d’amont est donc la rampe vestibulaire (RV) ; celle d’aval la rampe tympanique (RT) et entre les deux se trouve la rampe cochléaire (RC). La partie droite de la fig 3 correspond à une extrapolation verticalisée des trois rampes, comme si elles étaient dépliées en ligne droite avec à la base la position de l’étrier et de la fenêtre ronde : les deux rampes, vestibulaire et tympanique communiquent bien tout à fait au sommet de la cochlée en un point appelé hélicotréma (HT) ; cette communication permet aux ondes de parcourir les deux rampes, ondes qui prennent fin lors de la butée sur la fenêtre ronde ; mais dans la réalité le trajet complet est assez long et ne convient qu’à des trains d’onde de basse fréquence ; le plus souvent et d’autant plus que les sons sont aigus (haute fréquence) l’onde incidente s’interrompt là où la rampe cochléaire, qui est déformable, présente son point de résonance maximale ; ce point étant d’autant plus près de la base du colimaçon que le son est aigu ; l’onde est alors transmise à la rampe tympanique par la déformation de la rampe cochléaire sans passage par l’hélicotréma ce qui écourte le temps entre deux ondes successives (ce mécanisme est montré sur le schéma verticalisé sous la forme d’une ondulation de la rampe cochléaire qui permet la transmission de l’onde incidente vers la rampe tympanique sans avoir besoin de passer par le sommet) ; les sons perçus par l’oreille humaine se situent entre des fréquences de 20 hertz (sons graves) à 20.000 hertz (sons aigus) chez un sujet jeune bien entendant. L’intensité d’un son est aussi très importante ; elle se note en décibels (dB), chaque fréquence ayant sa limite de perception en fonction de l’intensité du son ; chez l’homme ce sont les sons graves entre 1000 et 3000 Hz qui nécessitent le moins d’intensité pour être perçus ; cette fréquence correspond à celle de la voix humaine ; c’est la raison aussi pour laquelle les sons graves de forte intensité (explosion) ont plus de risque de léser l’audition que les sons aigus ; avec l’âge la sensibilité aux sons aigus diminue nécessitant d’en augmenter l’intensité (presbyacousie). Le niveau de 1000 Hz est indiqué de manière très approximative pour montrer que la zone de meilleure vibration pour la réception de la voie humaine se trouve assez haut située dans la cochlée.
La fig 3 montre également le départ en arrière du labyrinthe osseux des deux nerfs qui forment le nerf auditif maintenant appelé plus judicieusement nerf vestibulo-cochléaire regroupant le nerf vestibulaire (NV) et le nerf cochléaire (NC) seul dédié à l’audition.
fig 4

On entre ici dans une spire sectionnée verticalement selon la ligne en pointillés verts de la fig 3. On retrouve les 3 rampes ; la rampe cochléaire (RC) contient l’organe sensoriel ou organe de Corti (OC) qui transforme les ondes reçues en influx nerveux ; elle est limitée en haut par une membrane très fine : la membrane de Reissner (MR) qui sépare donc la rampe cochléaire de la rampe vestibulaire et qui épouse toutes les ondes qui passent par cette dernière ; en bas la membrane basilaire (MB) faite de fibres à la fois raides et élastiques sépare la rampe cochléaire de la rampe tympanique : c’est elle qui vibre à la fréquence hertzienne des ondes incidentes, ses capacités de vibration maximale étant différentes du bas au sommet de l’hélice en raison d’une longueur et d’une souplesse différente de ses fibres tout au long de son trajet ; l’organe de Corti est composé de 4 rangées de cellules sensorielles reposant sur la membrane basilaire et se terminant par des cils ; lors de la vibration de la membrane les cils frottent contre une membrane assez épaisse située immédiatement au dessus, la membrane tectoriale (MT) ; quand les cils sont brossés dans un sens ils déclenchent un potentiel de dépolarisation qui est transmis à une fibre nerveuse sous-jacente qui représente la dendrite de la cellule excitée ; le noyau de cette cellule se trouve dans le ganglion spiral (GS) inclus dans l’os de la cochlée (en noir) ; de là l’axone rejoint le nerf cochléaire (NC). Dendrite, ganglion, axone et partie initiale du nerf se trouvent donc enchâssés dans l’os de la cochlée. Lorsque les cils sont brossés en sens inverse la cellule sensorielle bloque toute transmission d’influx.
Sur ces 4 rangées cellulaires seule la première (c1) est responsable de la transmission nerveuse principale ; les trois autres c2, c3 et c4 jouent sans doute un rôle modulateur qui n’est pas encore complètement éclairci.
Enfin il faut signaler la très grande fragilité des cils des cellules de l’organe de Corti pouvant être détruits définitivement par un son intense et la non possibilité de remplacement des cellules altérées par les cellules voisines contrairement aux cellules du goût et de l’odorat.
Tous les axones issus des ganglions spiraux se réunissent donc sur un tronc commun : le nerf cochléaire (NC) qui sort au centre de la face postérieure de la cochlée.
B – Du nerf cochléaire au cortex auditif
Le nerf cochléaire s’unit au nerf vestibulaire avant son entrée dans la boite crânienne, expliquant le terme ancien de nerf auditif pour désigner l’ensemble d’apparence unique ; mais il ne s’agit que d’une apparence chacun retrouvant sa voie particulière après l’entrée dans le tronc cérébral ; on continue néanmoins à donner un chiffre unique à l’ensemble soit la huitième paire (VIII) des nerfs crâniens.
La fig 1 montre la sortie du VIII du rocher et son entrée dans la boite crânienne à travers un conduit qui y est creusé : le conduit auditif interne (CAI) ; dans ce conduit il est accompagné du nerf facial (VII) en rouge pour le différencier et en raison de son activité motrice majeure pour les muscles du visage (le nerf facial est en effet surtout moteur : il sort de la boite crânienne par ce même orifice et suivra un trajet qui n’est pas traité ici – v, page sur la motricité du visage -).
Le VIII pénètre alors à la partie haute et latérale du bulbe (fig 5) juste au dessous du VII (v.page sur les nerfs crâniens) ; les deux composants vestibulaire et cochléaire se séparent immédiatement, chacun allant vers ses propres noyaux ; le nerf cochléaire (NC) se divise lui même en deux contingents qui se terminent sur trois noyaux cochléaires situés de part et d’autre du pédoncule cérébral inférieur (pci) : 2 noyaux antérieurs (NCA) et 1 postérieur (NCP).
fig 5

La fig 5 est une vue simplifiée du trajet intracrânien des fibres auditives ; le tronc cérébral est vu par sa face postérieure: le labyrinthe gauche (en bleu) est donc à gauche ; le droit (en orange) est à droite ; cette distinction a été faite pour mieux montrer le mixage des fibres D et G jusqu’au cortex auditif.
A partir des noyaux cochléaires les neurones de deuxième ordre montent vers les noyaux olivaires supérieurs (NOS) situés à la partie moyenne de la protubérance ; au cours de ce trajet la majorité des fibres croisent la ligne médiane ; la minorité reste homolatérale ; les NOS joueraient un rôle important dans nos facultés à localiser l’origine d’un son ; de chaque NOS les fibres de troisième ordre montent de façon homolatérale dans le faisceau lemniscal latéral (FLL) qui conduit au colliculus inférieur (CI) ; puis celles de quatrième ordre dans le corps genouillé médian (CGM) dépendance du thalamus ; celles de cinquième ordre se dirigent vers le cortex auditif primaire (CAP) en passant dans la substance blanche sous-lenticulaire (sbsl). Dans ce trajet à étapes des échanges de côté restent encore possibles, notamment entre les NOS et les CI. Le nombre de fibres montantes se restreint au fur et à mesure de cette montée, celles qui s’interrompent jouant probablement un rôle inhibiteur du type inhibition rétrograde sur certaines fibres montantes et permettant une sélection de la transmission pour les plus représentatives en éliminant soit les bruits de fond, soit les redondances gênantes pour une audition claire ; à signaler que certaines fibres nerveuses sont descendantes pour éviter également ce phénomène ; leur mauvais fonctionnement pourrait être en partie cause de certains acouphènes.
Le terme cortical final est la première circonvolution temporale dans sa partie supérieure (T1) ; il s’étend aussi en profondeur au cortex insulaire ; la fig 5 montre que chaque aire auditive reçoit des messages des deux oreilles avec une prévalence pour l’oreille controlatérale.

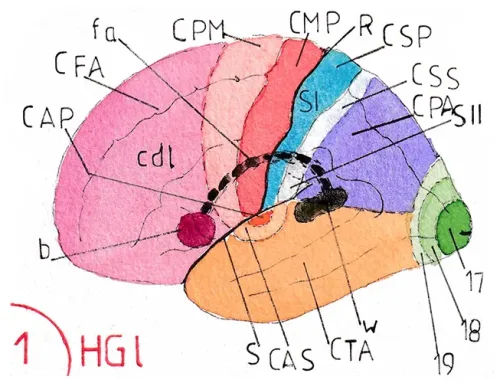
La fig 6 montre la face latérale de l’hémisphère gauche choisi ici en raison de son implication majeure dans la compréhension du langage grâce à l’aire de Wernicke (W) indiqué par une flèche, mais le processus est identique à droite ; la zone en bleu foncée correspondant à l’aire auditive primaire (CAP) ; en bleu clair à l’aire auditive secondaire (CAS) où commence une première interprétation des sons avant un traitement cortical complexe (v.page agnosie auditive ; partie neuropsychologique).
LF : lobe frontal ; LP : lobe pariétal ; LO : lobe occipital ; LT : lobe temporal.