Encéphale et motricité • 5 • Moelle épinière et
encéphale
Description anatomique
La moelle épinière (fig A , B et C) est partie intégrante du système nerveux central. A ce titre elle est une sorte de prolongement de l’encéphale en dehors de la boite crânienne ; comme lui elle est entourée de liquide céphalo-rachidien (LCR) qui circule librement d’une structure à l’autre. Les nerfs qui en sont issus sortent du canal osseux médullaire par les foramen intervertébraux et forment l’essentiel du système nerveux périphérique ; la fig A montre un fragment de moelle avec sa substance grise (sg) centrale en forme d’aile de papillon et la substance blanche (sb) alentour ; les ailes antérieures les plus larges sont le siège des corps cellulaires des neurones moteurs formant les racines motrices (rm) (en rouge) et les ailes postérieures, plus grêles, reçoivent les axones des nerfs sensitifs formant les racines sensitives (rs) (en bleu) ; la réunion des deux branches forment un nerf rachidien (nr) qui se divise à sa sortie de la colonne vertébrale en une branche antérieure (ba) (la plus volumineuse) destinée à la partie antérieure du corps et une branche postérieure (bp) à sa partie postérieure (viscères exclus).
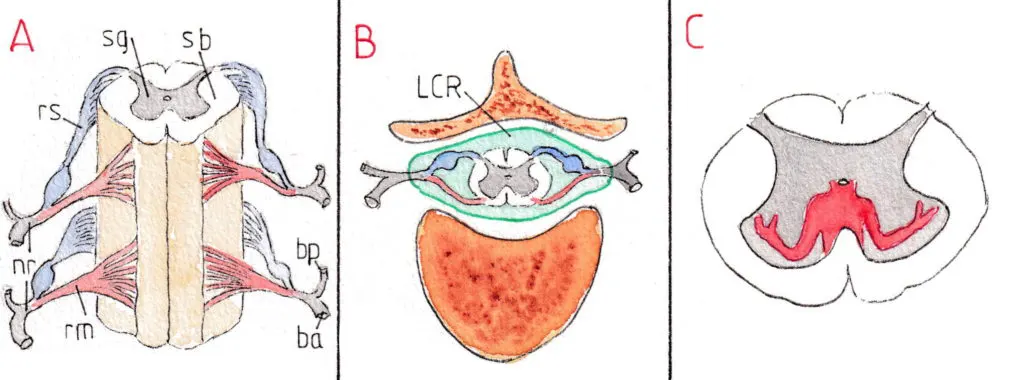
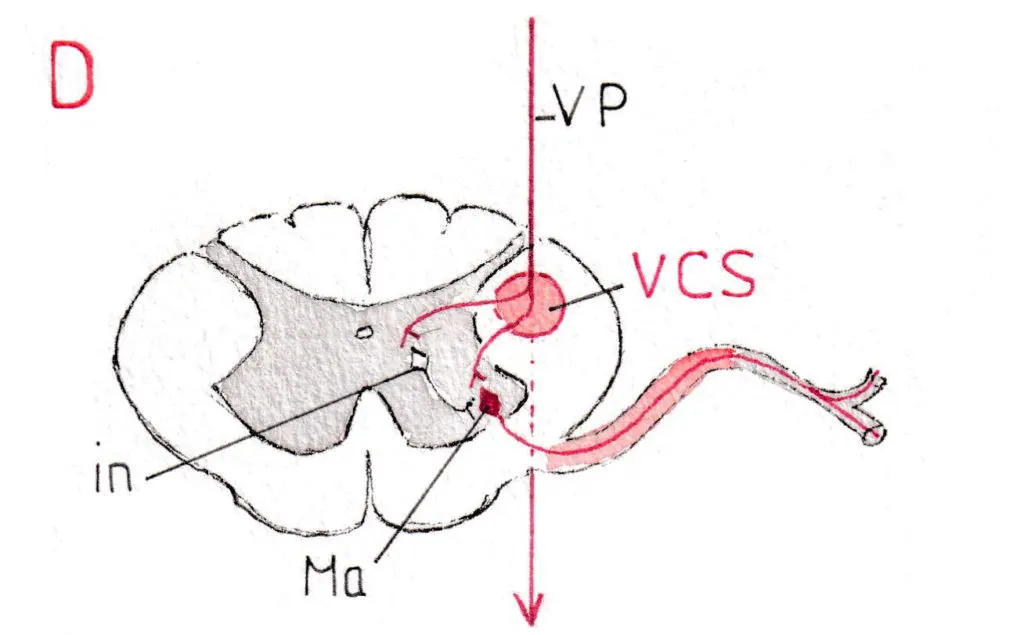
Les neurones moteurs sont appelés motoneurones alpha (Ma) et gamma (Mg) ; leur différence sera étudiée plus loin ; leurs corps cellulaires se trouvent dans les cornes antérieures (fig D) et se répartissent de façon systématisée en fonction de leurs destinations musculaires (fig C) ; ceux qui innervent les muscles les plus proximaux (axiaux et des ceintures) sont proches de la ligne médiane ; ceux qui innervent les plus distaux (mains ,doigts, pieds) sont les plus latéraux.
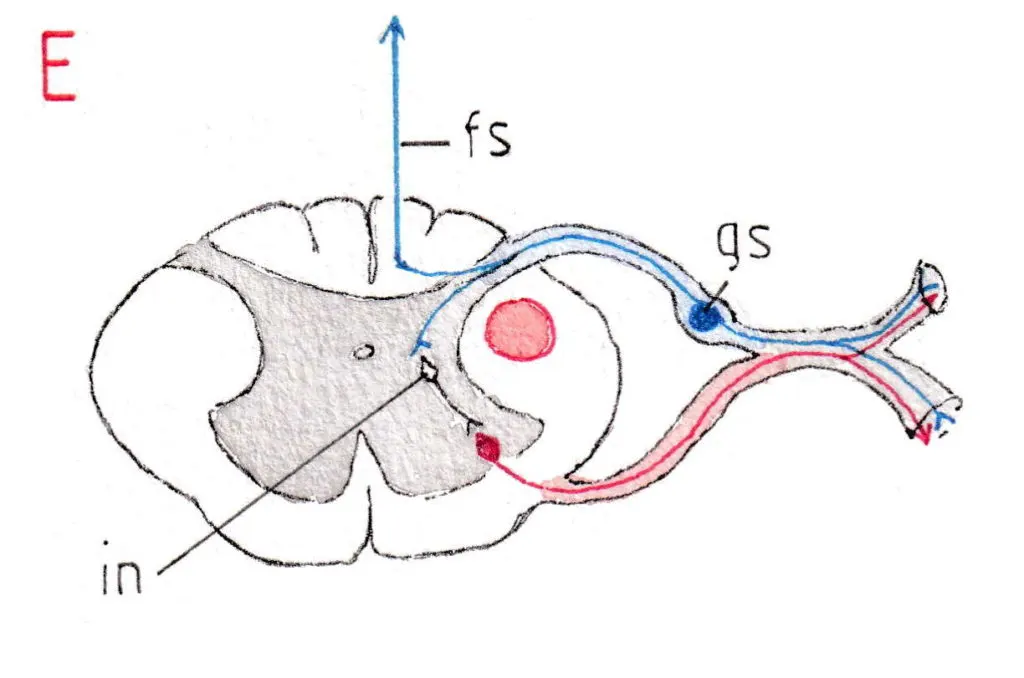
Les neurones sensitifs (fig .E) ont leurs corps cellulaires dans la racine sensitive où ils forment une sorte de renflement appelé ganglion spinal (gs) ; leurs axones pénètrent la moelle au niveau de la corne postérieure et se dirigent alors dans les faisceaux sensitifs (fs) remontant vers l’encéphale.
Description physiologique
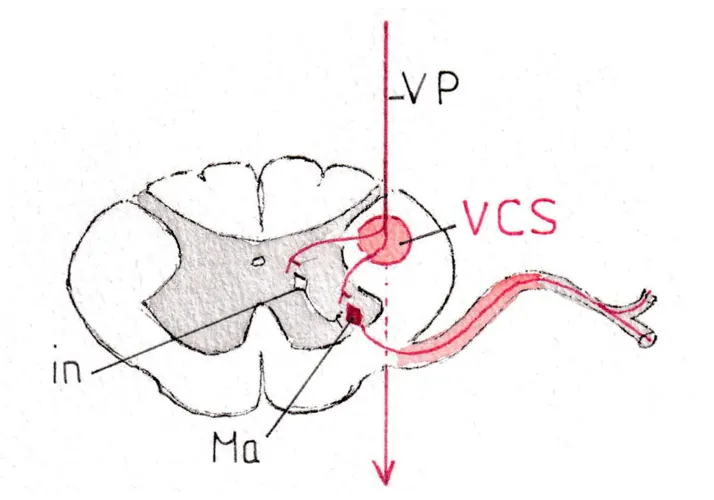
La fig D montre les connexions entre les neurones de la voie pyramidale (VP) qui après avoir été très antérieure dans le tronc cérébral est devenue postérieure et latérale dans la moelle, presque au contact de la corne grise postérieure et y prend le nom de voie cortico-spinale (vcs) ; la connexion se fait soit directement avec le corps cellulaire du motoneurone , soit le plus souvent par l’intermédiaire de courts neurones (interneurones -in-) siégeant dans la substance grise médullaire ; on peut suivre sur la fig D l’axone du motoneurone alpha (Ma) dans la racine antérieure et le début du nerf rachidien. Les motoneurones alpha sont donc sous la dépendance de la voie pyramidale volontaire issue du cortex moteur, dont ils constituent le prolongement vers les muscles concernés.
La Fig E met en jeu un neurone sensitif du même nerf rachidien ; il ramène l’information sensitive de la périphérie ; son corps cellulaire se trouve comme signalé plus haut dans un ganglion spinal (gs) de la racine postérieure; cette racine gagne la corne postérieure de la moelle puis l’axone du neurone sensitif gagne l’encéphale via les faisceaux sensitifs (fs) ; pendant sa traversée de la corne postérieure il envoie des prolongements courts vers le motoneurone le plus souvent avec le concours de neurones intermédiaires (interneurones -in-) certains pouvant être excitateurs, d’autres inhibiteurs ; cela montre comment sensitivité et motricité sont liées à l’étage médullaire pour y produire une automaticité qui peut souvent se passer du contrôle central.
Cette automaticité est favorisée par deux systèmes essentiels : le fuseau neuro-musculaire et l’organe de Golgi :
On entend par fuseau neuro-musculaire -fnm- (fig F et F’) la dispersion à l’intérieur des muscles de la motricité volontaire d’un certain nombre de petites structures musculo-sensorielles sous forme de fuseaux séparés , mais faisant corps avec les fibres musculaires adjacentes et ayant une fonction sensitive très particulière ; la fig F montre un de ces fuseaux dessiné à un fort grossissement pour mieux montrer sa structure ; le schéma est très simplifié pour faciliter la compréhension du fonctionnement du fuseau :
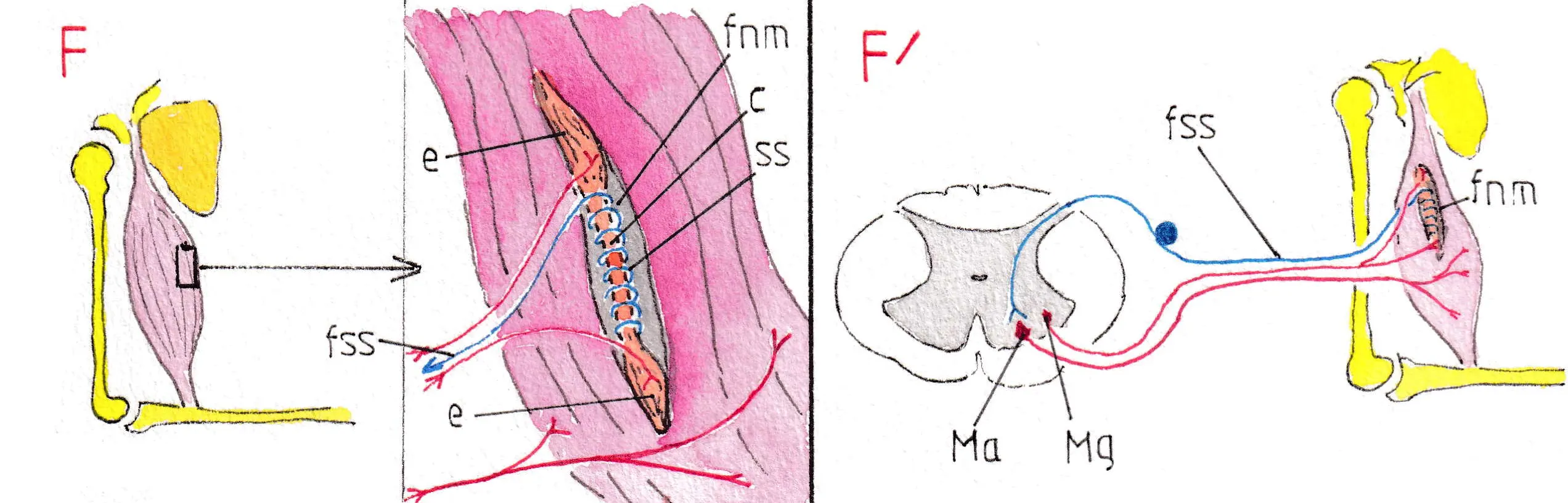
ce dernier est en effet formé de deux extrémités (e) faites de fibres musculaires contractiles et d’un centre (c) non contractile entouré par une spirale sensitive (ss) qui peut transmettre des informations à la moelle selon son degré d’étirement ; ces influx sensitifs sont transportés par une fibre sensitive spécialisée (fss) à très haute vitesse de conduction et qui suit le trajet des autres fibres sensitives via les racines sensitives postérieures ; par ailleurs les parties contractiles du fuseau sont sous la dépendance d’un motoneurone de la corne antérieure de la moelle appelé , comme signalé plus haut, motoneurone gamma (Mg) (fig F’) ; l’influx qui vient de la voie pyramidale atteint en réalité les deux motoneurones (alpha et gamma ) en même temps, entrainant une contraction simultanée des fibres musculaires et de celles du fuseau, de telle sorte que la spirale sensitive reste sous une tension constante ; mais si le muscle se trouve davantage sollicité par un effort supplémentaire (par exemple un poids à soulever) les fibres musculaires vont avoir tendance à s’allonger, de même que le fuseau qui leur est contigu ; la distension de la spirale sensitive autour de la zone non contractile produit une volée d’influx dans le nerf sensitif spécialisé, lequel est en connexion excitatrice directe avec le motoneurone alpha correspondant ; ce dernier augmente alors la puissance de ses influx vers le muscle pour le faire contracter davantage et ainsi maintenir l’effort supplémentaire nécessaire (fig F’).
L’arrêt volontaire de la contraction musculaire désamorce le système. Ce mécanisme peut ainsi être assimilé à une sorte de servo -assistance car elle se passe à l’échelon local et n’oblige pas le cortex à agir de lui-même ; l’état de réaction du fuseau neuro-musculaire est souvent utilisé en clinique par la recherche du réflexe d’étirement dit réflexe myotatique : ainsi quand on frappe le tendon rotulien avec un marteau à reflexe on étire les fibres musculaires du muscle antérieur de la cuisse et l’on provoque , en raison de l’étirement imprévu du fuseau une volée d’influx par sa fibre sensitive spécialisée ; laquelle , via le motoneurone alpha va faire contracter le muscle pour raccourcir ses fibres.
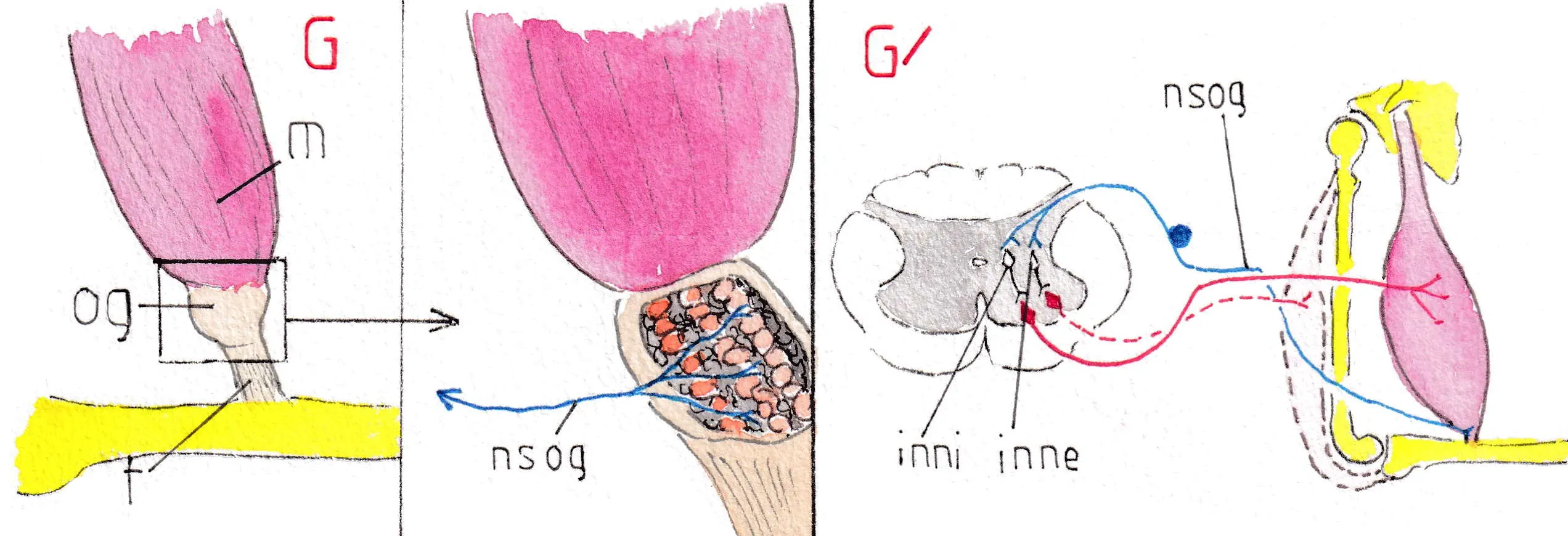
L’organe de Golgi (og ) (fig G et G’) complète ce dispositif local en inhibant les trop fortes tensions sur les tendons qui pourraient courir un risque d’étirement traumatique en cas d’effort musculaire excessif :
l’organe de Golgi de situe entre muscle (m) et tendon (t) ; il est fait de fibrilles entrelacées au sein desquelles les terminaisons sensitives se trouvent plus ou moins comprimées suivant l’état de tension du tendon ; ces terminaisons appartiennent à des fibres sensitives de l’organe de Golgi (nsog) différentes de celles des fuseaux mais également spécialisées ; elles gagnent la corne postérieure par la voie sensitive commune et elles entrent en connexion dans la substance grise de la moelle avec un neurone inhibiteur (inni) qui diminue les décharges du motoneurone alpha et donc oblige à un relâchement momentané du muscle ; si le relâchement est suffisant pour diminuer la traction sur le tendon , les influx sensitifs venant de l’organe de Golgi s’arrêtent et les influx activateurs du motoneurone alpha reprennent ; il s’agit là d’un mécanisme de protection musculo-tendineux également automatisé sur le principe du servo- contrôle local ; par ailleurs si le relâchement musculaire n’est pas assez rapide la fibre sensitive issue de l’appareil de Golgi peut même activer un muscle antagoniste par l’intermédiaire d’un interneurone excitateur (inne) (ici en pointillé un extenseur de l’avant bras sur le bras ).
Ensemble comparatif des schémas moelle epinière 1
La moelle épinière a donc surtout une fonction motrice subordonnée à l’encéphale ; mais elle a aussi des fonctions motrices autonomisées ; il en est ainsi de réactions de défense en flexion ou extension en cas d’excitation douloureuse cutanée et cela de la façon qui parait la plus logique à la protection du corps ; de très nombreuses expériences chez l’animal, quadrupède en particulier, ont pu montrer le rôle médullaire prépondérant pour les mouvements de locomotion que ce soit en marche normale ou au galop ; il est vraisemblable que l’automaticité médullaire joue un rôle très important également dans l’enchainement des mouvements de la marche et de la course chez l’homme une fois ceux-ci initiées par la commande corticale.
Dans les séquelles motrices des cérébrolésions l’apparition d’une spasticité caractérisée par une hypertonie musculaire atteint souvent les muscles paralysés, manifestations aggravant les séquelles de la paralysie ; deux mécanismes semblent prédominants : d’une part la libération excessive des voies réticulospinales (voir voies réticulaires et motricité), normalement excitatrices mais soumises à une inhibition corticale qui disparait ou diminue dans les cérébrolésions ce qui favorise l’excitation de la substance réticulée sous-jacente. Ce mécanisme serait particulièrement responsable des hypertonies précoces contemporaines de l’éveil et associées à des phénomènes neurovégétatifs plus ou moins sévères : hypersudation, hypertension artérielle, troubles respiratoires avec hypersécrétion bronchique ; l’hypertonie est le plus souvent diffuse avec membres supérieurs en hyperflexion et membres inférieurs en hyperextension ; elle est souvent transitoire et s’atténue alors au bout de 2 à 3 mois.
Les hypertonies secondaires apparaissant plusieurs semaines après la cérébrolésion ; elles seraient davantage lièes à un emballement du reflexe myotatique comme expliqué plus haut, chaque relâchement musculaire spontané étant contré par la boucle des fuseaux neuromusculaires ; quand elle est limitée à quelques muscles c’est une très bonne indication à l’injection de toxine botulique directement dans le muscle pour bloquer le récepteur musculaire de l’acetylcholine neuromédiateur des terminaisons musculaires des axones des motoneurones alpha ; quand elle est plus diffuse l’indication d’injection dans le liquide céphalorachidien perimédullaire d’un agoniste( activateur) d’un inhibiteur naturel (le GABA) de la transmission nerveuse permet de diminuer à la fois l’hyperexcitabilité des fuseaux neuro-musculaires ainsi que les réactions neuro-végétatives générales ; dans certains cas très sélectionnés on peut avoir recours à des sections partielles de nerfs moteurs (et uniquement eux) pour diminuer l’activité du réflexe myotatique responsable.